

Abb. 1: Spitzahorn am Frauenberg südwestlich der Stadt Grafenau (Bayerischer Wald). Foto: Foto: Bernhard Rau (AWG)
Drei der fünf Acer-Arten kommen in Deutschland häufiger vor: der Bergahorn, der Spitzahorn und der Feldahorn. Bei der Trockenheitstoleranz nimmt der Spitzahorn eine Zwischenposition ein zwischen Berg- und Feldahorn. In einem Gewächshausexperiment weisen gerade die jungen Sämlinge eine hohe Trockenheitstoleranz auf. Als Baumart der Ebene und der mittleren Gebirgslagen ist die Prognose im Klimawandel recht unterschiedlich: Im Alpenvorland und im Jura zeichnet sich ein geringes Anbaurisiko ab; auf der warmen und trockenen Fränkischen Platte nimmt es zu. Als typische Mischbaumart ist der Spitzahorn auf mittleren bis gut Nährstoff versorgten Böden einzeln oder truppweise beteiligt. Das natürliche Verbreitungsgebiet erstreckt sich von Griechenland bis zum Ural in Russland. Potenziell kommt der Spitzahorn in ganz Bayern vor. In höheren Anteilen ist er in den Wäldern Unter- und Mittelfrankens beteiligt, was sich auch in der Verteilung der zugelassenen Erntebestände und der gemeldeten Populationen widerspiegelt.
Das Anpassungspotenzial wurde bisher nicht detaillierter untersucht. Die Ausweisung von Saatguterntebeständen erfolgte ausschließlich anhand des Phänotyps. Wie bei anderen Nebenbaumarten sind beim Spitzahorn geringe Mindestkriterien (Mindestalter: 40 Jahre, keine Mindestfläche, Mindestbaumzahl: Im Bestand 20, für die Ernte 10) vorgegeben.
Im Rahmen des Projekts "AcCarTi" werden die Herkunftsempfehlungen und Herkunftsgebiete für die Baumarten Spitzahorn (Acer platanoides L.), Hainbuche (Carpinus betulus L.) und Sommerlinde (Tilia platyphyllos Scop.) auf genetischer Grundlage überarbeitet, um die Erntebasis zur Deckung des zukünftigen Bedarfs an Vermehrungsgut zu verbessern.
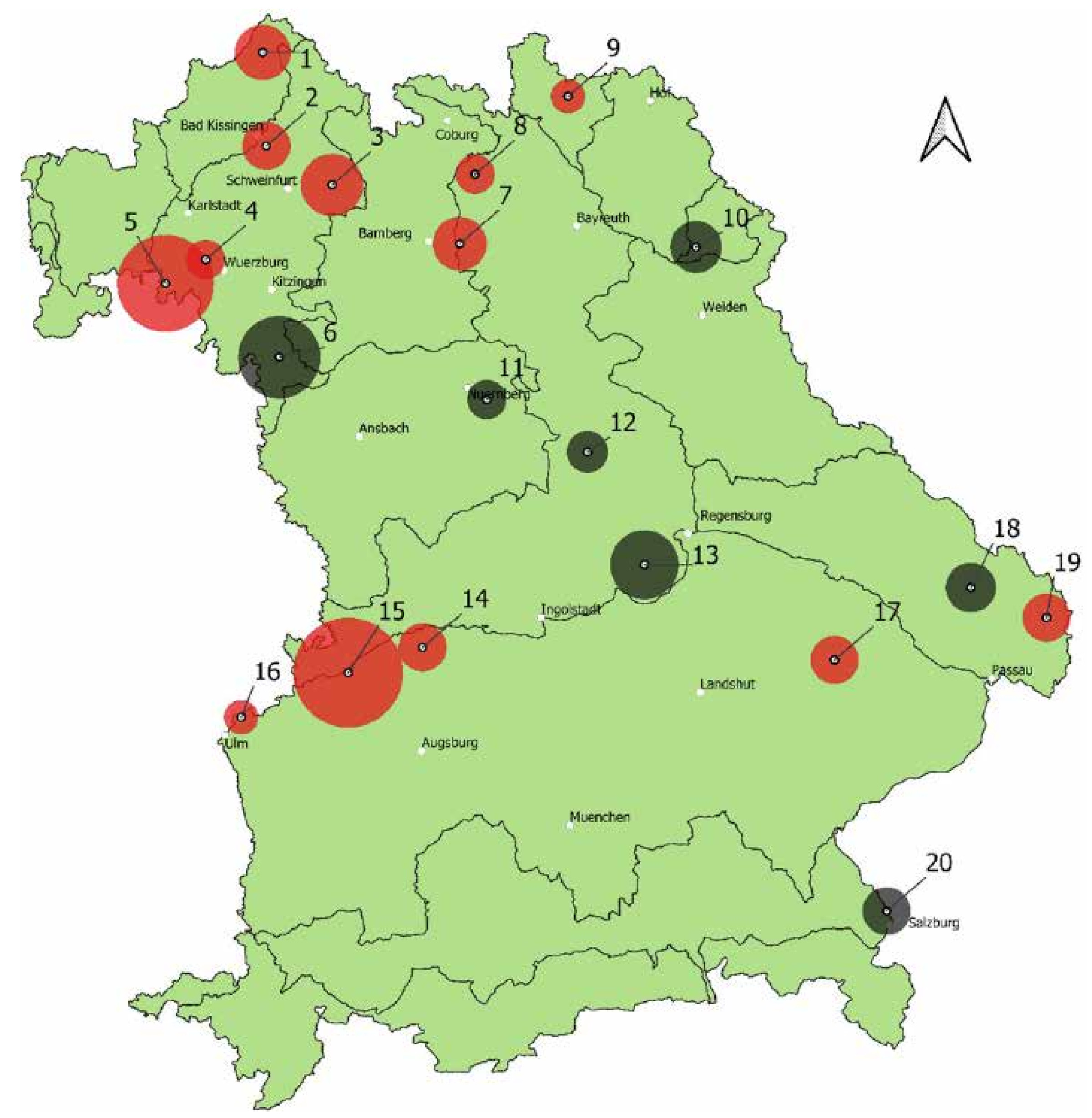
Abb. 2: Karte der 20 beprobten Bestände des Spitzahorns (rot = EZR, grün = neue Bestände, die nicht zugelassen sind). Die Größe der Kreise stellt die Größe der Population dar (Anzahl der Spitzahorne).
Zu Beginn des Projekts wurde für jede Baumart ein Ist-Zustand erhoben. Gleichzeitig erfolgte eine Abfrage der Ämter für Ernährung, Landwirtschaft und Forsten (ÄELF) und der Bayerischen Staatsforsten (BaySF) mit der Bitte um Rückmeldung von potenziellen Vorkommen. Diese sollten bestimmte Voraussetzungen (z. B. Mindestbaumzahl und -alter, Qualität, Isolierung und Autochthonie, Fruktifikationsfähigkeit) erfüllen. In Bayern gibt es aktuell (2021) 29 zugelassene Erntebestände für Spitzahorn, die sich folgendermaßen aufteilen: Zwei Bestände im Herkunftsgebiet (HKG) 800 03 und 27 im HKG 800 04. Von diesen 29 Beständen wurden 25 begangen und schließlich 13 beprobt (Nr. 1, 2, 3, 4, 5, 7, 8, 9, 14, 15, 16, 17, 19, darunter zwei DKV-Bestände: Nr. 2 und 16). Die BaySF meldeten 52 Vorkommen, dazu kamen sieben Meldungen über die ÄELFs. Aus diesen wurden sieben Vorkommen ausgewählt und beprobt (Nr. 6, 10, 11, 12, 13, 18, 20). Südlich der Donau gab es kaum noch Meldungen von größeren Vorkommen. Die Verteilung der untersuchten Populationen zeigt die Karte in Abb. 2.

Abb. 3: Bei einer konsequenten Pflege sind stattliche Dimensionen von bis zu 80 cm BHD möglich. Foto: Gregor Aas
In den Beständen kommt der Spitzahorn mit einem Anteil von 1 bis 20 % vor. Er ist meist einzeln bis truppweise beigemischt. Auf den trockeneren Standorten oder wenn die Buche in ihrer Konkurrenzkraft nachlässt, ist er in höheren Anteilen vertreten und auch in Gruppengröße beteiligt. An vielen Standorten tritt er häufiger an Bestandesrändern und entlang der Wege auf. Einige der Vorkommen sind aus Pflanzungen (ca. in den 1960er-Jahren) entstanden und haben sich gut entwickelt. Dort wurde der Spitzahorn gerne zusammen mit Bergahorn gepflanzt. Wenn ein Nebenbestand vorhanden war, zeigten die Bäume immer eine bessere Qualität (Stammform, Beastung). In einigen Vorkommen wurde die Pflege vernachlässigt oder Durchforstungen zurückgestellt, was in der Qualität und Vitalität sichtbar wird. Die Spitzahorne wachsen zwar mit, entwickeln aber eingezwängte, kleine Kronen und schlechte Stammformen (krumm, schief, schwache Dimension). Wenn auf eine ordentliche Entwicklung geachtet wurde, werden Baumhöhen von 34 m und ein BHD bis zu 80 cm erreicht.
Auf den von ihm bevorzugten Standorten tritt der Spitzahorn oft in Konkurrenz zu Bergahorn, Esche und Buche. In trockeneren Bereichen kommt er auch noch zusammen mit Feldahorn, Elsbeere oder Speierling vor.
In fünf beprobten Beständen konnten offensichtlich Symptome der Rußrindenkrankheit an Bergahornen festgestellt werden. Nebenstehende Spitz- oder Feldahorne zeigten bisher in den allermeisten Fällen keine Anzeichen. Dies könnte auf eine geringere Anfälligkeit des Spitzahorns hindeuten, der als hitze- und trockentoleranter gilt als der Bergahorn. Trotzdem sollte die Entwickelung der Rindenkrankheit weiterhin genau beobachtet werden.
Genetische Analysen und Methoden
Die für DNA-Analysen verwendeten Mikrosatelliten sind kurze, nicht kodierende DNA-Abschnitte, die Wiederholungen von zwei (z.B. TA) oder drei (z.B. GAC) Basen enthalten. Für die genetischen Analysen beim Spitzahorn wurden neue Marker im Auftrag von Dr. Heino Konrad, österreichisches Bundesforschungs- und Ausbildungszentrum für Wald (BFW) in Wien, im Microsynth ecogenics Labor (Schweiz) entwickelt und für diese Untersuchung freigegeben. Nach der ersten Analyse der genetischen Daten und der Bewertung der Marker wurden jedoch zwei der 15 Marker von der weiteren Analyse ausgeschlossen. Somit wurde die gesamte Analyse auf der Grundlage von 13 Markern durchgeführt. Ausgehend von den für die Einzelbäume bestimmten Multilocus-Genotypen wurden Allelhäufigkeiten und Parameter berechnet, die die genetische Variation innerhalb der Vorkommen beschreiben.
Ergebnisse der genetischen Analysen an Spitzahorn
Die Anzahl der Allele (Na) bei den 20 untersuchten Spitzahorn-Vorkommen schwankte zwischen 5,92 (Population 12-Velburg) und 9,31 (Population 15- Steinheim) mit einem Mittelwert über alle Vorkommen von Na = 7,72. Für die Ausweisung von Saatguterntebeständen wird die mittlere Anzahl an effektiven Allelen Ne herangezogen. Dieser Wert steht für die genetische Diversität innerhalb einer Population und bezieht die Häufigkeitsverteilung der Allele mit ein. Die Anzahl der effektiven Allele (Ne) schwankte zwischen 3,34 (Population 12 – Velburg) und 5,13 (Population 11 – Fischbach) mit einem Mittelwert von Ne = 4,29. Die Heterozygotie (He) war in allen untersuchten Populationen nicht sehr hoch (Mittelwert = 0,738). Die allelische Vielfalt (Ar) schwankte zwischen 5,82 (Population 12 – Velburg) und 9,17 (Population 15 – Steinheim) mit einem Mittelwert von Ar = 7,63. Insgesamt waren 32 Privatallele vorhanden, die sich über 15 Populationen verteilten. Der Inzuchtkoeffizient (FIS) variierte von -0,078 (Population 12 – Velburg) bis 0,103 (Population 15 – Steinheim).

Abb. 4: Verteilung der genetischen Variationsparameter auf die Vorkommen
Die Analyse der molekularen Varianz (AMOVA) zeigte, dass 91 % der gesamten genetischen Variation innerhalb der Populationen und 4 % zwischen den Populationen vorkommt. Die Hauptkoordinatenanalyse (PCoA) zeigt die genetischen Hauptmuster, die in der genetischen Distanzmatrix nach Nei enthalten sind. Die Populationen 4, 12 und 13 liegen dabei am weitesten von den anderen Populationen entfernt.
Räumlich genetische Strukturen
Abbildung 5 zeigt das Ergebnis der Clusterbildung auf Basis der 13 eingesetzten Microsatelliten-Genorte. Die zwei (K = 2) und vier (K = 4) Farben geben die verschiedenen STRUCTURE-Cluster wieder. Jeder einzelne Baum ist durch eine vertikale Farblinie dargestellt. Die Länge einer jeden Linie (y-Achse) entspricht der Wahrscheinlichkeit der individuellen Zuteilung zu einem von STRUCTURE gebildeten Cluster.
Die Bayes‘sche Clusteranalyse mit der Software STRUCTURE 2.3.3 ergab eine mögliche Struktur von zwei und vier genetischen Clustern. Dabei wurde der größte Wert für deltaK bei K = 2 (deltaK = 12,64) erreicht und der nächst größere Wert für deltaK bei K = 4 (deltaK = 4,20). Bei zwei Clustern trennen sich die Populationen 4 und 12 am weitesten ab mit den größten Anteilen der jeweiligen Cluster. Die anderen Populationen zeigen eine ziemlich gleiche Verteilung der beiden Cluster. Bei vier Clustern trennt sich die Population 12 sehr eindeutig von den anderen Populationen ab. Die Bestände 18 und 19 werden von einem lila Cluster dominiert und der blaue Cluster ist eher im Westen vorherrschend. Insgesamt wurde eine Beimischung aller Cluster beobachtet.
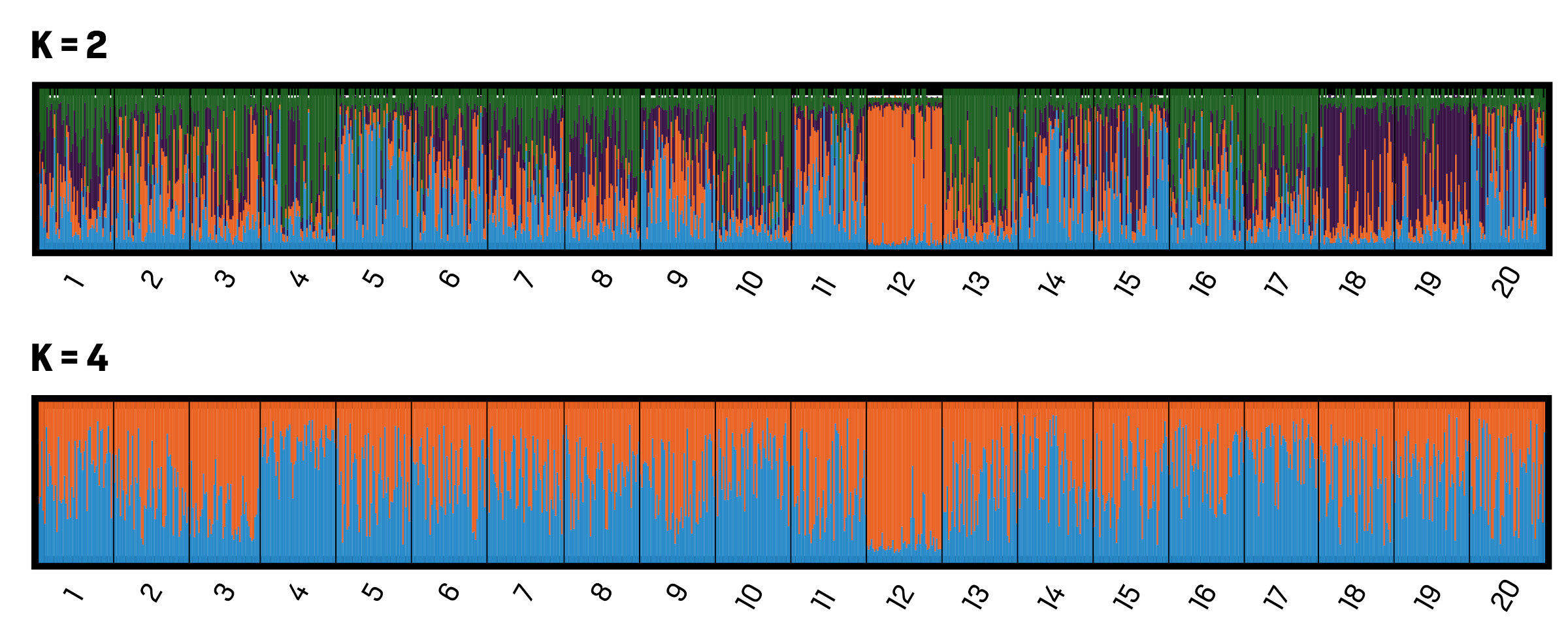
Abb. 5: Histogramm der genetischen Strukturen der 20 Spitzahornvorkommen in Bayern.
Diskussion und Schlussfolgerung
Baumarten wie Spitzahorn haben bisher eine untergeordnete Rolle gespielt oder wurden als dienende Baumarten genutzt, weshalb deren Anpassungspotenzial bisher nicht detaillierter untersucht worden ist. Die hier erfolgte erstmalige genetische Untersuchung des Spitzahorns in Bayern ermöglicht einen ersten Überblick über dessen genetische Variabilität. Dadurch können Empfehlungen für Maßnahmen zur Erhaltung und Nutzung dieser Baumart abgeleitet werden. Die genetische Vielfalt von Erntebeständen sollte möglichst hoch sein, um eine hohe Anpassungsfähigkeit des Saat- und Pflanzgutes zu gewährleisten. Nach den vorliegenden Ergebnissen kann eine geringe genetische Differenzierung zwischen den Spitzahornbeständen beobachtet werden.

Abb. 6: Verteilung der räumlich-genetischen Strukturen in Bayern anhand der ökologischen Grundeinheiten. Die Farbverteilungen K1 bis K4 zeigen keine klare räumliche Gruppierung an.
Die Verteilung der räumlichgenetischen Strukturen in Bayern (Farbverteilungen K1 bis K4) zeigt einzelne räumliche Gruppierungen an. Im HKG 800 03 ist eine Durchmischung zu beobachten. Diese Tatsache kann dadurch erklärt werden, dass bisher nur zwei Saatguterntebestände (9 und 19) zugelassen waren und Saatguterntebestände aus dem HKG 800 04 als Ersatzherkunft genutzt werden konnten. Aus den Ergebnissen lässt sich ableiten, dass 19 der untersuchten Bestände als mögliche Saatgutquelle empfohlen werden. Sieben Bestände wurden im Rahmen des Projekts neu gemeldet und untersucht. Die empfohlenen Bestände sollten eine hohe genetische Diversität sowie eine hohe allelische Vielfalt ausweisen. Der Bestand 12 Velburg wird nicht zur Beerntung vorgeschlagen, da dieser die geringsten Vielfaltsparameter aufweist und aus Vermehrungsgut von wenigen Elternbäumen entstanden sein könnte. Bei dem Bestand 4 Margetshoechheim ist die genetischen Diversität Ne = 3,4 gering. Die allelische Vielfalt Ar = 6,82 ist mit anderen Beständen vergleichbar.
Bei einer günstigen Kombination der genetischen Diversität Ne und der allelischen Vielfalt Ar kann bei einer Auswahl von mindestens 30 Bäumen, die über das ganze Vorkommen verteilt sind, eine Saatguternte durchgeführt werden. Es wird empfohlen, bevorzugt Saatguterntebestände auf einer größeren Fläche (min. 0,5 ha reduziert) und einer guten räumlichen Verteilung im Bestand auszuweisen. Bei Spitzahorn ist durch das FoVG keine Mindestfläche vorgegeben. Es kann beobachtet werden, dass in der Regel bei allen größeren Beständen die genetischen Vielfaltsparameter überdurchschnittlich abschneiden. Des Weiteren sollte die Anzahl der zu beerntenden Bäume bei Nebenbaumarten erhöht werden. Gerade bei Nebenbaumarten, bei denen keine großen Bestände vorhanden sind und der Genfluss zwischen den Beständen kaum vorhanden ist, sollten mindestens 30 Bäume einer Waldpopulation beerntet werden.

Abb. 7: Blätter des Spitzahorns. Foto: Christoph Josten
Für die Erhaltung forstlicher Genressourcen beim Spitzahorn werden in allen relevanten Generhaltungszonen Erhaltungsbestände ausgewiesen. Erhaltungsmaßnahmen bei Waldbäumen sollten sich auf die Erhaltung der genetischen Variation innerhalb der Art konzentrieren, aber auch die Einzigartigkeit des bestehenden Genpools berücksichtigen. Bestände, die eine allelische Vielfalt über Ar > 7,6 aufweisen, werden in erster Linie als Erhaltungsbestände empfohlen. Gerade bei seltenen Baumarten ist der Aufbau von Erhaltungsplantagen eine wichtige Maßnahme und kann zur Sicherung forstlicher Genressourcen und zur Versorgung mit hochwertigem Vermehrungsgut beitragen. Im Rahmen des Projekts konnten insgesamt 58 Plusbäume aufgenommen werden, die zur Anlage einer Erhaltungsplantage dienen können. Bei Baumarten wie dem Spitzahorn, die im Klimawandel an Bedeutung gewinnen und eine positive Prognose aufweisen, wäre es sehr wichtig, zeitnah eine neue Erhaltungsplantage anzulegen.
Die Förderung des Projektes erfolgt durch das Bayerische Staatsministerium für Ernährung, Landwirtschaft und Forsten (StMELF) über das Kuratorium für forstliche Forschung.




