
Im Vergleich zur Landwirtschaft scheint in der Forstwirtschaft die Handlungsoption "Anpassung durch den Austausch von genetischem Material" nahezu irrelevant. Lange Lebenszyklen, in weiten Teilen kaum domestizierte Arten und späte Reproduktionsfähigkeit machen Waldbäume als "genetische Objekte" eher unattraktiv. Dennoch ist die Herkunftsforschung in der Forstwissenschaft ein langjähriges und etabliertes Feld, welchem im Klimawandel wieder eine stärkere Bedeutung zukommen muss.
Natürliche genetische Anpassung oder "assisted migration"?
Häufig liest man im Zusammenhang mit genetischer Anpassung an den Klimawandel auch den englischen Begriff "assisted migration". Dieser Begriff subsumiert jedoch verschiedene Formen der anthropogen gesteuerten Migration von Baumarten (siehe Kasten). Die von uns nun beschriebene Handlungsoption der genetischen Anpassung ist ein Teilaspekt der "assisted migration", und zwar der sogenannte "assisted gene flow".
Die gedankliche Grundlage dieser Handlungsoption ist, dass sich bei unseren heimischen Baumarten infolge vielfältiger Umwelteinflüsse in den letzten Jahrtausenden eine große genetische Variation ausbilden konnte. Ursache dafür sind die Prozesse:
- Mutation
- Selektion
- Genfluss (im Zuge der Reproduktion) und
- genetische Drift.
Für eine verbesserte lokale Anpassung sind wesentlich die Mutation und die Selektion verantwortlich. Die Selektion, die beispielsweise bei der Naturverjüngung von Wäldern zum Tragen kommt, kann nur die Eigenschaften selektieren, die schon im genetischen Portfolio einer Population (Bestäubungseinheit) vorhanden sind. In dem sich natürlicherweise immer die Baumindividuen vermehren, die am besten mit den vorherrschenden Standortsbedingungen zurechtkommen, ergibt sich eine gerichtete Anpassung an den Standort. Die Mutation verläuft hingegen ungerichtet. Es werden Erbinformationen falsch kopiert, wodurch in der nächsten Generation tatsächlich neue Eigenschaften entstehen. Allerdings wirken diese Eigenschaften häufig zulasten der Baumfitness und nur selten kommt es zu einer vorteilhaften Mutation.
Beide Prozesse führen dazu, dass sich eine Baumart in einem Gebiet über mehrere Generationen immer mehr an die lokalen Gegebenheiten anpasst. Es entstehen die sogenannten Populationen oder in der Forstsprache Herkünfte bzw. Provenienzen. Die Umweltbedingungen im Herkunftsgebiet sind somit eng verknüpft mit den lokal angepassten Populationen.
Assisted migration
Es lassen sich 3 Typen von "assisted migration" unterscheiden:
- Herkunftstransfer (assisted population migration, assisted gene flow): Populationsverbringung, bei der Saatgut einer Baumart innerhalb ihres derzeitigen Verbreitungsgebiets klimatisch oder geografisch verlagert wird. Dieses geschieht in der Regel über Herkunftsgebietsgrenzen hinweg: z. B. Verlagerung der Buche 200 km nach Norden innerhalb ihres Verbreitungsgebiets.
- Erweiterung/Verschiebung des Verbreitungsgebiets (assisted range expansion): Saatgut wird aus dem aktuellen Verbreitungsgebiet in angrenzende, für die Baumart geeignete Gebiete verbracht. Es findet eine Erweiterung des Verbreitungsgebiets statt: z. B. Anbau von Elsbeere in Mecklenburg-Vorpommern, die bislang nur weiter südlich vorkommt.
- Anbau nichtheimischer Baumarten (assisted species migration): Hierbei werden Baumarten von weit außerhalb ihres derzeitigen Verbreitungsgebiets angesiedelt: z. B. Japanische Lärche in Mecklenburg- Vorpommern. Dieses kann aus wirtschaftlichen Gesichtspunkten, aber auch aus Gründen der Erhaltung geschehen, wenn die Art in ihrem natürlichen Verbreitungsgebiet gefährdet ist.
"Local is best!"
Was passiert jedoch, wenn sich die lokalen Bedingungen rasant ändern? Die plakative Aussage "Local is best!" ist spätestens dann zu hinterfragen. Nützt der Baumart dann noch die jahrhundertelange lokale Anpassung an die bislang herrschenden Standortsbedingungen? Und was ist unter dem Begriff "lokal" überhaupt zu verstehen?
"Lokal" wird bei der In-situ-Erhaltung forstlicher Genressourcen sehr eng ausgelegt. Man bezieht sich dabei auf Einzelbäume oder einen geschlossenen Bestand, in dem man die Naturverjüngung oder das Saatgut nutzt. Hier kann man sicher sein, dass tatsächlich auch die lokal vorhandene genetische Information an die nächste Generation übergeben wird.
In der waldbaulichen Praxis legt man den Begriff "lokal" deutlich weiträumiger aus. Die Grundlage bilden die Herkunftsgebiete. Herkunftsgebiete sind zumeist großräumige Areale, die ein oder mehrere Wuchsgebiete überspannen. Sie werden entsprechend der natürlichen Differenzierung einer Baumart ausgewiesen, z. B. Herkunftsgebiet 810 02 der Rotbuche – "Ostsee-Küstenraum". Die entsprechenden rechtlichen Regelungen finden sich im Forstvermehrungsgutgesetz (FoVG) und in den Herkunftsgebietsempfehlungen für M-V.
Mit diesen geografischen Abgrenzungen sind jedoch noch keine Baumeigenschaften (Überlebensfähigkeit, Wachstum, Resistenz, etc.) definiert. Vielmehr sind die Übergänge zwischen den Gebieten fließend und auch innerhalb eines Herkunftsgebietes gibt es selbstverständlich genetische Unterschiede. Einerseits sind viele Bestände aufgrund der fast völligen Entwaldung Europas gar nicht autochthon. Andererseits bilden sich auch innerhalb von Herkunftsgebieten mehrere Populationen heraus. Diese Bestände kennen wir zumeist in Forstkreisen, wenn sie durch überragende Wuchseigenschaften und markige Namen Bekanntheit erlangt haben (z. B. Güstrower Kiefer, Selber Höhenkiefer, Schlitzer Lärche, Kiekindemarker Douglasie). Wie gut sind aber nun die lokalen Populationen an die aktuellen Bedingungen angepasst und inwieweit können sie mit sich ändernden Standortsbedingungen umgehen?
"Genetische" Versuchsflächen in M-V
Für diese Fragestellungen werden Versuchsflächen der Forstgenetik / Forstpflanzenzüchtung angelegt. Auch in Mecklenburg-Vorpommern existieren zahlreiche dieser Flächen (Abb. 1). Die Flächen kann man in drei Typen unterteilen: 1) Herkunftsversuche (gelb), 2) Nachkommenschaftsprüfungen (blau), und 3) Klonprüfungen (grün).
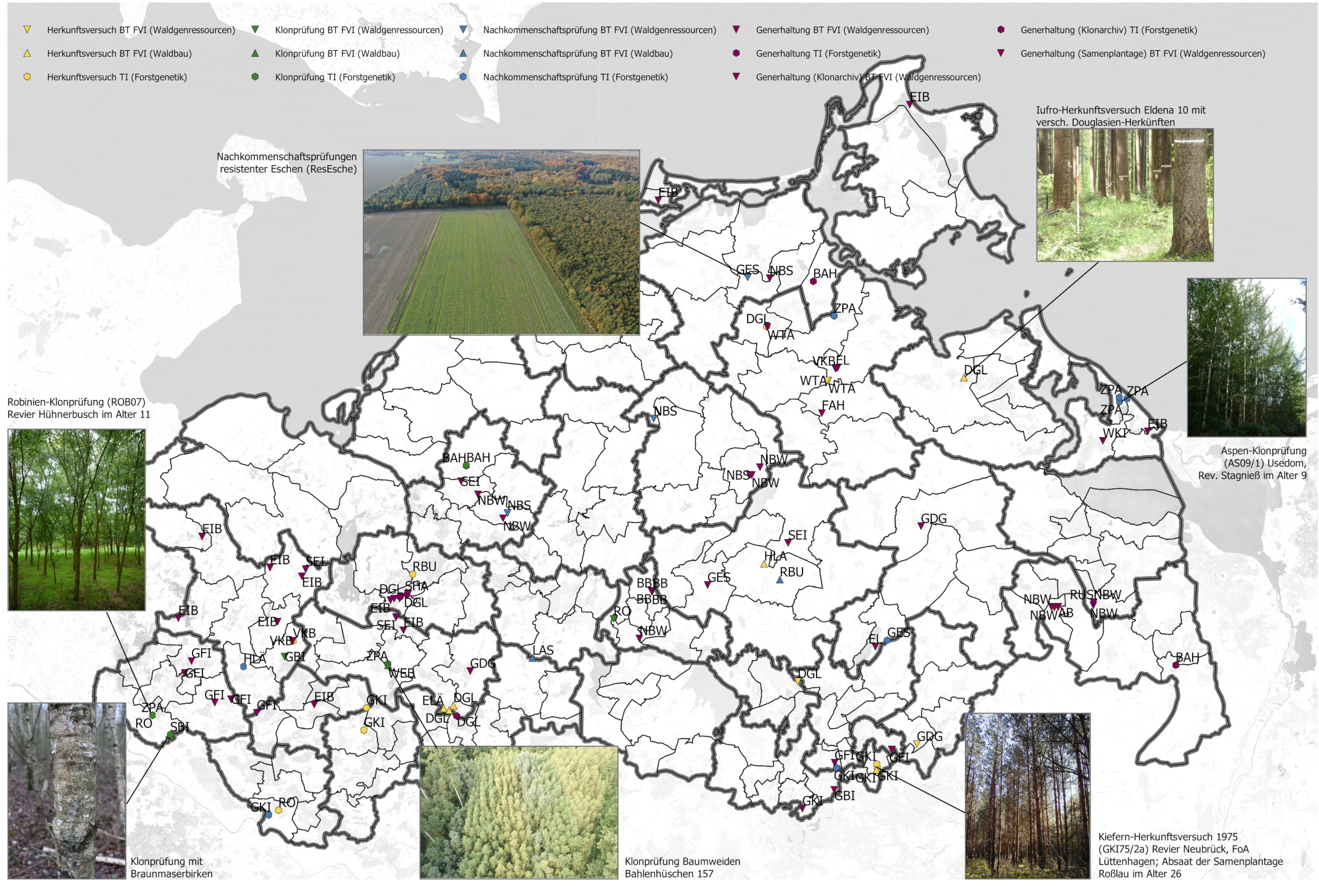
Abb. 1: Alle genetischen Versuchsflächen und eine Auswahl wichtiger Generhaltungsbestände in den jeweiligen Forstämtern/-revieren. Neben den genetischen Versuchsflächen im Land gibt es noch weitere Arten von langfristigen Versuchsflächen, die im ForstGaia hinterlegt sind. Die Kürzel benennen die Baumart. Die Symbole und deren Farben repräsentieren die Versuchsart und das verantwortliche Institut. Bei Fragen zu Versuchsflächen in M-V können Sie sich an das Forstliche Versuchswesen wenden.
Herkunftsversuchen
Die bekanntesten Flächen sind sicherlich die Herkunftsversuche. Aus dem gesamten Verbreitungsgebiet werden hier Herkünfte an einem Standort zusammengebracht, um deren unterschiedliche Eigenschaften unter gleichen Standortsbedingungen zu vergleichen. Die größten Herkunftsversuche im Land sind zwei Douglasie-IUFRO - Versuche aus den 70-er Jahren (Eldena 10 und Parchim 21). Sie beinhalten Douglasien aus 26 unterschiedlichen Herkunftsgebieten Nordamerikas und M-V und sind jeweils rund 9 ha groß. Die Versuche werden auch heute noch regelmäßig aufgemessen.
Besonders bei den Herkunftsversuchen wird deutlich, wie stark die lokale Anpassung ausgeprägt ist, und wie viel "assited gene flow" tatsächlich bringt. Ein Herkunftsversuch der Waldkiefer (siehe Abb. 2) zeigt, dass südwestliche Herkünfte (z. B. Lampertheim, Südhessen) unter den hiesigen Klimabindungen (Klimastufe t) im Schnitt 15 % bis zu 29 % mehr Holzzuwachs leisten als die lokale Herkunft. Ein Saatgutbestand aus Polen (Supraśl) bringt es sogar auf 39 %! Eine unfassbare Leistung, wenn man bedenkt, dass die Bonität direkt gekoppelt ist mit den Bodenertragswerten.
Dennoch ist der überragende Wachstumsgewinn mit Vorsicht zu genießen. Wie es die "Super"-Baumart nicht gibt, so existiert auch nicht die "Super"- Herkunft. Aus Herkunftsversuchen der Douglasie ist bekannt, dass Herkünfte mit hohen Wuchseigenschaften eine geringere Trockenstressresilienz besitzen können. Man erkauft sich also in solch einem Fall höhere Produktivität zulasten von geringer Stabilität.
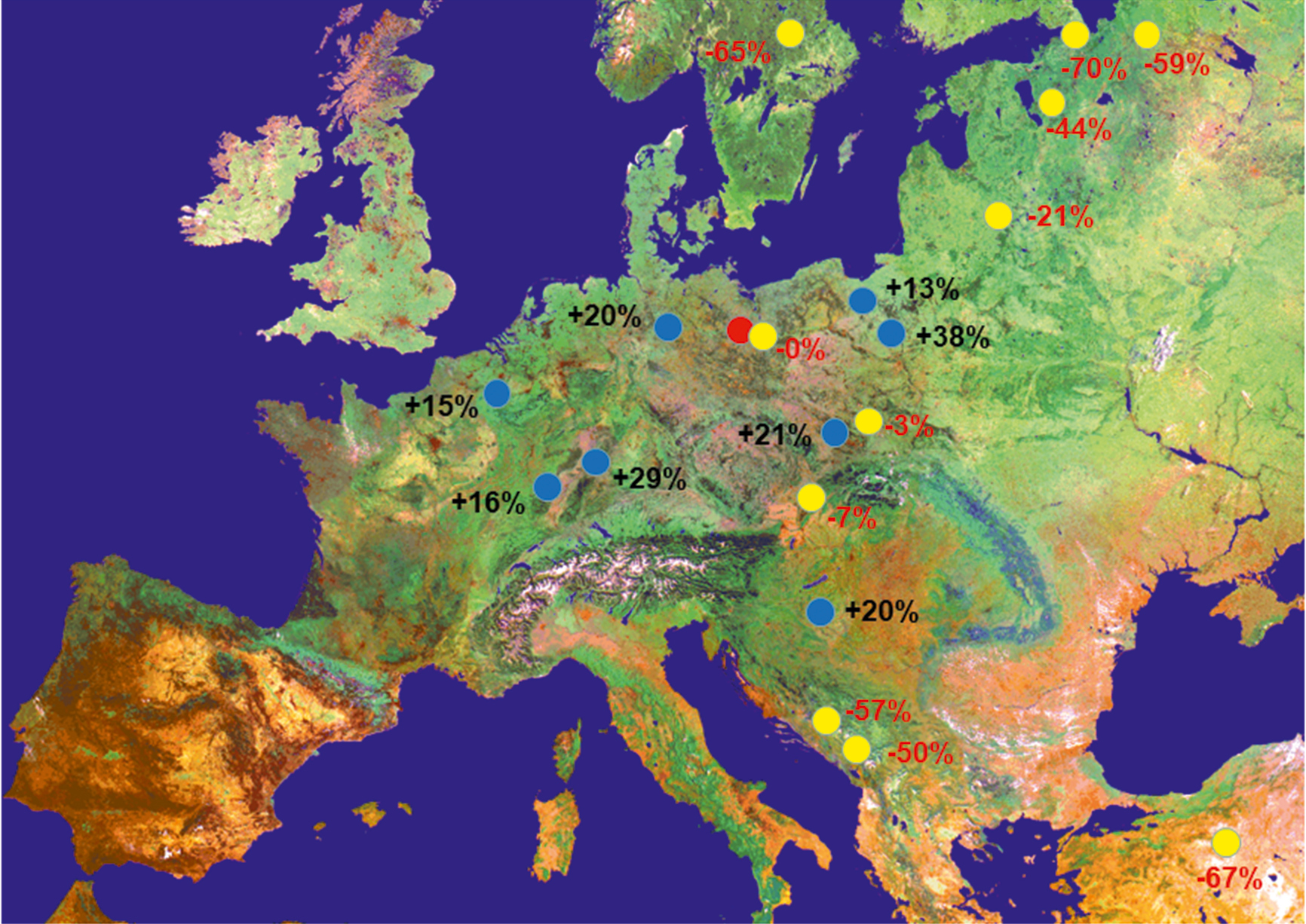
Abb. 2: Örtlich muss nicht das Beste sein – Unterschiede im Schaftholzvolumen in einem 28-jährigen Kiefernversuch in Ostbrandenburg (rot: örtliche, blau: bessere, gelb: schlechtere Herkünfte)

Abb. 3: Hier wachsen seit 30 Jahren Kiefern aus Mittel- und Osteuropa nebeneinander – Dreenkrögen 52. Foto: E. Thurm
Nachkommenschaftsprüfungen
Die Nachkommenschaftsprüfungen sind ein weiterer Bestandteil der genetischen Forschung und ein zentrales Element der Forstpflanzenzüchtung. Hier wird nicht auf das Herkunftsgebiet geschaut, sondern auf die Anlagen von Einzelbäumen. Sie können als Leistungsprüfungen der Nachkommen oder zur Analyse quantitativ genetische Parameter (Heritabilität, Kombinationseignung, Zuchtwert) genutzt werden. Die Erzeugung des Versuchsmaterials kann dabei durch gelenkte Kreuzungen oder freie Abblüte der entsprechenden Auslesebäume erfolgen.
Eine große Nachkommenschaftsprüfung befindet sich u. a. im Forstamt Schuenhagen. Hier werden gegen das Eschentriebsterben resistente Eschenindividuen untersucht. Auch bei der Baumart Kiefer gibt es Flächen zweier Nachkommenschaftsprüfungen in M-V. Im Revier Drewin wurde 1990 eine Fläche einer Prüfung mit 81 Nachkommenschaften aus kontrollierten Kreuzungen angelegt (zwei weitere Flächen in Brandenburg). Nach 31 Jahren konnten Nachkommenschaften mit überdurchschnittlicher Wuchs- und Qualitätsleistung identifiziert werden. So übertrafen die besten Nachkommenschaften die Absaat einer Samenplantage (Waldsieversdorf-Güstrow) um 7,5 – 13,3 % im Höhenwachstum. Ebenfalls konnten Elternbäume mit positiven Zuchtwerten für dieses Merkmal identifiziert werden. Weitere Nachkommenschaftsprüfungen in M-V finden sich zur Schwarznuss, mit dem Ziel standortsangepasste Individuen dieser nordamerikanischen Baumarten zu detektieren (Abb. 4).
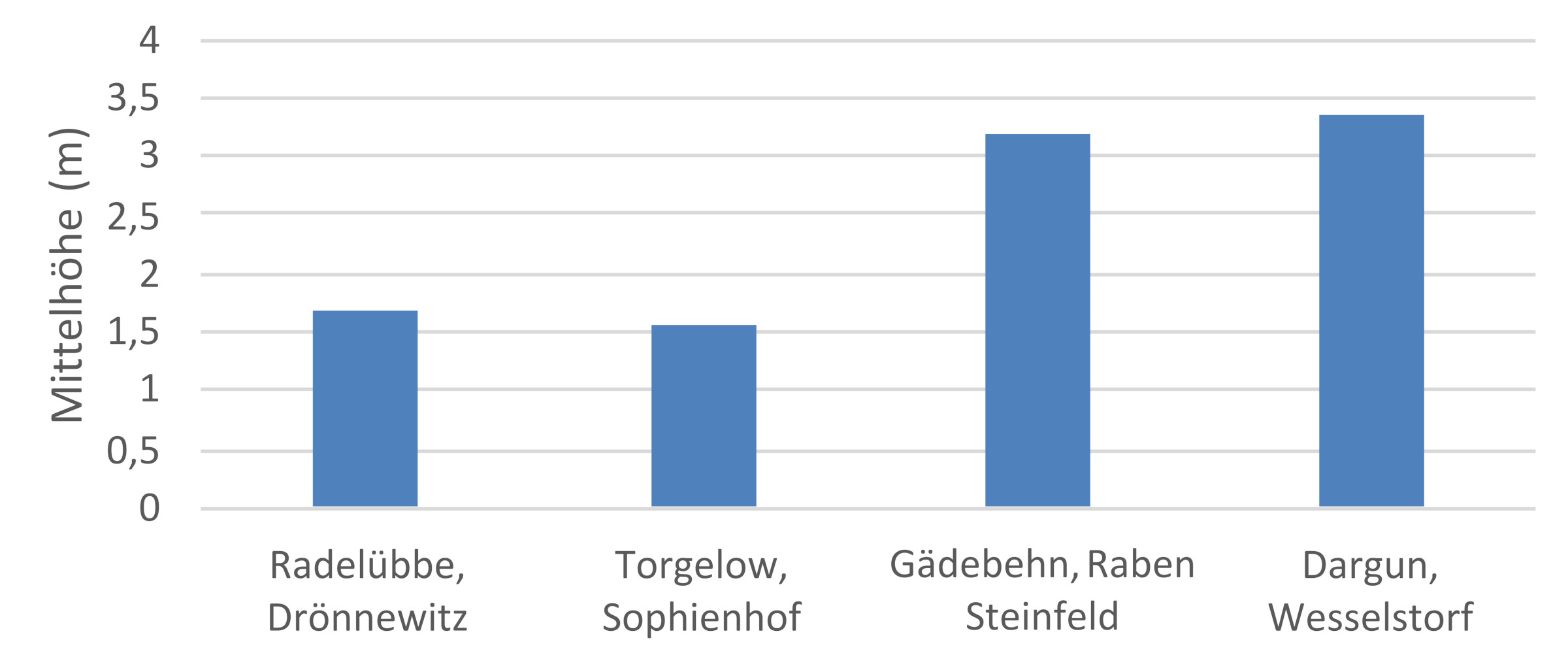
Abb. 4: Höhenwuchsleistung von 7-jähriger Schwarznuss Absaaten auf einer Versuchsfläche im FoA Schlemmin. Bei den Herkünften Raben Steinfeld und Wesselstorf beträgt sie die doppelte Höhe der beiden anderen Absaaten Rade-Jübbe und Torgelow.
Klonprüfungen
Klonprüfungen bilden eine weitere Kategorie von forstgenetischen Versuchen. Hier werden durch vegetative Vermehrung (Gewebekultur, Stecklingsvermehrung, Pfropfung) gewonnene erbgleiche Abkömmlinge verschiedener Einzel-(Plus-)bäume zum Vergleich ausgepflanzt. Bei der Anpassung an den Klimawandel hat dieses Verfahren den Vorteil, schnell auf spezielle Eigenschaften wie Trockenstresstoleranz selektieren zu können. Vegetative Vermehrung führt jedoch zu keiner genetischen Weiterentwicklung (Evolution). Beispiel für Klonprüfungen in M-V sind die Versuchsfläche Hühnerbusch mit Robinie oder Besitz mit Birke (beide im Forstamt Schildfeld). Bei der Robinie werden Klone geradschaftiger Ausgangsbäume geprüft. Bei der Birke sind es Klone mit speziellen Holzeigenschaften (Maserung).
Generhaltung in M-V
Neben den natürlichen Prozessen steuert auch der Mensch im erheblichen Maße die genetische Anpassung. Dennoch gibt es auch in der Genetik viele Unbekannte, und ein vermeintliches "Trockenstress"-Gen kann unter sich ändernden Umweltbedingungen ganz andere Reaktionen bewirken. Daher ist es wichtig, das genetische Potenzial, welches durch Adaption entstanden ist, zu sichern.
Dafür werden sogenannte Generhaltungsbestände ausgewiesen. Dabei unterscheidet man einerseits die In- situ-Generhaltung, wenn das Genmaterial am Ort (lokal) erhalten wird (Abb.1, lila Symbole). Ein Beispiel sind die Ivenacker Eichen. Andererseits gibt es die Ex-situ-Generhaltung, bei der erhaltenswerte forstliche Genressourcen ausgelagert werden, entweder Saatgut oder pflanzliches Gewebe in Genbanken, in Beständen oder Sammlungen abseits der Einsammlungsgebiete, die durch Saat, Pflanzung oder mit Klonen angelegt werden.
In der Landesforstanstalt wurden z. B. Klonarchive von Restvorkommen für die Elsbeere, die Eibe und die Europäische Schwarzpappel aufgebaut. Samenplantagen wurden zum Erhalt der Arten Holzapfel und Wildbirne angelegt. Diese Anlagen wurden stets durch genetische Untersuchungen begleitet, um die genetische Identität zu gewährleisten.
Genetische Vielfalt im Waldbau
Letztlich laufen zahlreiche Aktivitäten, um im Klimawandel sowohl das genetische Potenzial der Baumarten zu sichern als auch deren genetische Anpassung zu steuern. Wie weit kommen diese Aktivitäten jedoch zum Tragen, wenn das Ziel Dauerwald heißt?
Der Dauerwald wird im Wesentlichen getragen von der Idee der großflächigen natürlichen Verjüngung. Das größte Potenzial, die natürliche genetische Vielfalt zu erhalten, wird dabei in der artenreichen und kontinuierlichen Naturverjüngung gesehen. Auch in den Leitlinien der Landesforstanstalt M-V soll die natürliche Verjüngung den Regelfall der Walderneuerung darstellen.
Die von uns genannten Informationen zur genetischen Vielfalt widersprechen diesem Konzept nicht. Denn trotz der Rasanz des Klimawandels muss die Ausnutzung natürlicher – wenn auch sehr langsamer – Anpassungsprozesse gewahrt bleiben. Am wichtigsten ist es daher, sich als Waldbewirtschafter*in bewusst zu machen, dass man mit seinem Handeln im Wald einen Einfluss auf die genetische Vielfalt hat. Dabei kann sie in verschiedenen Behandlungsphasen unterschiedlich beeinflusst werden. Kritisch zu beurteilen ist es, wenn Naturverjüngung nach einer längeren Phase der Zielstärkennutzung eingeleitet wird.
Hier sind die besten Vererber zum Zeitpunkt der Verjüngung bereits nicht mehr vorhanden. Zielstärkennutzung vor Beginn der natürlichen Verjüngung bedeutet Selektion auf negative Merkmale.
Eine zu drastische Stammzahlreduktion in der Durchforstungsphase führt zur Einengung des genetischen Materials. Die Auslesedurchforstung ist deutlich besser geeignet, denn sie greift der natürlichen Selbstdifferenzierung vorweg. Bei der Hiebsart erwirken langfristige und kleinflächige Verjüngungsmethoden eine höhere genetische Diversität. In den untersuchten Beständen war die Wahrscheinlichkeit erhöht, dass möglichst viele Bäume ihre Gene in die nächste Generation weitertragen. Der Großschirmschlag erwies sich in der gleichen Studie als nachteilig, da bei den wenigen verbleibenden Individuen ein hoher Anteil Selbstbefruchtung beobachtet werden konnte. Letztlich sollte jedoch auch hier nicht zu rigoros auf nur eine Hiebsart gesetzt werden. Le Lefèvre (2004) betont in seiner großen Literaturauswertung, dass eine Vielfalt von Hiebsarten ebenfalls zur genetischen Vielfalt beiträgt. Besonders Bestände mit einem Überhang von schlecht veranlagten Bäumen könnten hier von einer menschlich gesteuerten Selektion profitieren (Abb. 5).

Abb. 5: Buchenbestand mit schlechten Stammformen und hohem Zwieselanteil, was sich auch in der Naturverjüngung wiederfindet. Foto: M. Liesebach
Der größte Anteil der Beeinflussung des genetischen Materials erfolgt jedoch bei der Kunstverjüngung. Besonders aussichtsreich kann "assisted gene flow" bei Arten sein, die über bekanntermaßen hohe genotypische Unterschiede bzw. deutlich unterschiedliche Herkünfte verfügen. Dazu zählen beispielsweise die Waldkiefer, Gewöhnliche Fichte, aber auch die Douglasie.
Bei der Buche ist die Notwendigkeit von "assisted gene flow" keinesfalls eindeutig. Aktuelle Untersuchungen aus dem Projekt "BuVit MV" zeigten zwar, dass gerade nicht autochtone Buchenherkünfte in Mecklenburg-Vorpommern Tendenzen zur höherer Dürreanfälligkeit besaßen. Jedoch ist diese Variation zwischen den Populationen scheinbar geringer als die Varianz innerhalb der Populationen. Eine extreme Option der genetischen Anpassung könnten hier Hybridisierungen mit der trockentoleranteren Orientbuche darstellen, die jedoch erst in der Folgegeneration auftreten würden.
Eine weitere Option der Bewirtschafter*innen, positiv auf die genetische Vielfalt bei der Kunstverjüngung einzuwirken, ist es Einheitssortimente zu vermeiden. Konnert et al. (2007) konnten nachweisen, dass bei Buche, Fichte und Tanne die großen Sortimente in der Baumschule die niedrigste genetische Vielfalt aufwiesen. Bei der Etablierung einer Baumart ist auch die Mischung von Saatgut aus unterschiedlichen Herkunftsregionen eine Option sich breiter aufzustellen. Hier kann je nach erwarteter Klimaerwärmung und Risikoaffinität der Anteil trockenstresstoleranterer Pflanzen gegenüber lokalem Vermehrungsgut angepasst werden.
Fazit

Abb. 6: Das Vorhandensein vieler "Vererber" über lange Zeiträume ist besonders bei der Buche ein Garant für hohe genetische Vielfalt. Wichtig ist daher neben Qualitätskriterien auch Vitalitätskriterien in die Durchforstungsentscheidung einfließen zu lassen. Foto: E. Thurm
Am Ende möchten wir zu unserer provokativen Überschrift "Pflanzen wir Buchen unter Buchen?" zurückkommen. Eine Pflanzung von trockenstresstoleranten Rotbuchen in einem bestehenden Rotbuchenaltbestand ist letztlich keine sinnvolle Handlungsoption. Bei augenscheinlich nicht an den Klimawandel angepassten Buchenreinbeständen gilt es zuerst weitere Baumarten einzubringen, um über die Baumartenmischung eine Stabilität des Bestandes zu erreichen.
Genetische Anpassung über künstliche Verjüngung muss auf der Landschaftsebene gedacht und auf der Bestandesebene durchgeführt werden. Das bedeutet, die Bewirtschafter*innen ergänzen den Kiefernanteil in ihren Forstämtern bei der "Neuanlage" von Beständen um trockenstresstolerante Herkünfte. Ein Beispiel wären Aufforstungsflächen, bei denen keine entsprechenden Samenbäume vorhanden sind.
Eine weitere Erkenntnis aus unserem Beitrag ist, dass bei der genetischen Anpassung, wie schon bei der Einführung neuer Baumarten, ein erheblicher Forschungsbedarf besteht. Die Kernbotschaft an die Bewirtschafter*innen ist: „Seien Sie sich bewusst, dass Ihr waldbauliches Handeln die genetische Vielfalt des Waldes immer beeinflusst!“ Die waldbaulichen Eingriffe sollten die natürlichen Prozesse fördern, um die genetische Vielfalt zu erhalten und den Genfluss zu begünstigen. Dadurch wird lokale Anpassung weiterhin ermöglicht, sodass die heutigen Bestände und die ihnen zugewiesenen Waldfunktionen erhalten bleiben.



