
Einleitung
Der Speierling (Sorbus domestica L.) ist eine der seltensten Baumarten in Deutschland und erreicht in den Bundesländern Thüringen, Sachsen und Sachsen-Anhalt seine nördliche Verbreitungsgrenze in Mitteleuropa. Die Populationen hier dürften nach der letzten Eiszeit etwa zwischen 9.000 bis 8.000 v Chr. von Südwestfrankreich nach Deutschland über das Rhonetal und den Jura eingewandert sein (KAUSCH-BLECKEN VON SCHMELING 2000). Somit dürfte das Vorkommen der Art auch in Thüringen einen natürlichen Ursprung haben. Durch das sich ändernde Klima hat der Speierling in den letzten Jahren sicherlich an Interesse gewonnen, ist er doch sehr trockenresistent und bringt dennoch sehr wertvolles Holz, das auch beachtliche Dimensionen erreichen kann. Deswegen und aus Naturschutzgründen wurde die Art in den letzten Jahren auch relativ häufig angepflanzt.
Auch in den anderen Teilen Europas ist der Speierling selten, häufiger kommt er nur in naturbelassenen Trockenwäldern in Südfrankreich vor. Molekulargenetische Untersuchungen haben gezeigt, dass die Population großräumig in zwei Teilpopulationen geteilt ist: GEORGE et al. (2015) konnten zeigen, dass eine Populationsgruppe in Westeuropa existiert, eine weitere Populationsgruppe hat sich im östlichen Teil Europas gebildet. Beide Gruppen dürften sich aber im Mittelmeerraum mischen, da hier die größte Diversität gefunden wurde; dies ist auch Zeugnis der Nutzung durch den Menschen, die viele hunderte Jahre zurückgeht. Insbesondere während des Römischen Reiches dürfte viel Handel mit Früchten getrieben worden sein, sodass die Unterschiede in diesem Bereich relativ undeutlich sind.
In Thüringen wurden bisher 185 vermutlich autochthone Altspeierlinge vorgefunden. Speierlinge bilden keine Reinbestände, die Exemplare kommen meist in Einzelmischung vor. Die Schwerpunktvorkommen befinden sich in den Bereichen Grabfeld, Kyffhäuser, Hainleite und Saaletal auf kalkgeprägtem Grundgestein und haben dort überdauert, wo Nieder- und Mittelwaldwirtschaft am längsten betrieben wurde bzw. auf Standorten, wo die Buche konkurrenzschwach ist. Nur hier konnte sich der lichtbedürftige Speierling überhaupt langfristig in Einzelmischung etablieren.
In der vorliegenden Arbeit wurde zum ersten Mal die genetische Vielfalt der Thüringer Speierlinge untersucht. Als Vergleichsbasis dienten die Daten aus GEORGE et al. (2015), sodass die Proben auch mit dem gesamteuropäischen Muster der Populationsstruktur verglichen werden konnten. Zusätzlich wurde eine Reihe von Jungspeierlingen, die in der jüngeren Vergangenheit gepflanzt worden waren, untersucht. So sollte auch festgestellt werden, ob für die Aufforstungen heimisches Material verwendet worden ist.
Material und Methoden
Insgesamt wurden für die Untersuchung Blattproben von 66 Speierlingen (Sorbus domestica L.) aus Thüringen verwendet. Davon waren 41 Altspeierlinge aus verschiedenen Regionen Thüringens, die restlichen 25 Proben sind Jungspeierlinge, die zwischen 1995 und 2000 gepflanzt wurden.
Die eingelangten Blattproben von Sorbus domestica wurden zunächst zwischengelagert (Zugabe von Silicagel zur Trocknung, Lagerung bei -18°C). Anschließend wurden ca. jeweils 2 cm² Blattmaterial in Eppendorf-Mikrozentrifugenröhrchen überführt, in flüssigem Stickstoff schockgefroren. Anschließend wurde die DNA aus den Proben mittels des Qiagen DNeasy Kits extrahiert.
In Folge wurden die Proben an einem rps16-Chloroplastenmarker, sowie 7 nuklearen Mikrosatelliten mittels PCR wie in GEORGE et al. (2015) beschrieben genotypisiert. Die Größe der resultierenden DNA-Fragmente wurde auf einem CEQ8000 (Beckman-Coulter) mittels Kapillargelelektrophorese bestimmt.
Ergebnisse
Chloroplasten-Marker
Insgesamt konnten von den eingesandten 66 Proben bei 59 die Chloroplasten-Marker erfolgreich angewandt werden, bei 65 Proben konnten zumindest 5 der 7 Mikrosatelliten-Marker erfolgreich amplifiziert werden. Lediglich von Probe Nr. 61 (Jungspeierling) konnten keine Daten erlangt werden, hier war offenbar die Probenqualität nicht ausreichend für eine erfolgreiche DNA-Extraktion (trotz mehrmaligen Versuchen).
Die Analyse der Daten des Chloroplasten-Markers mittels der Software GENALEX (PEAKALL & SMOUSE 2012) zeigte bereits einen großen Unterschied in der genetischen Vielfalt und in der Haplotypen-Zusammensetzung der Stichproben. Die Ergebnisse sind in Tab. 1 zusammengefasst.
Tab. 1: Diversität von Sorbus domestica in Europa und in Thüringen: Anzahl Individuen (N); Anzahl Allele (Na); effektive Anzahl Allele (Ne); Information-Index (I); haploid diversity (h); unbiased haploid diversity (uh); F – Frankreich, CH – Schweiz, EB – Elba/Italien, CS – Korsika/Frankreich, IT – Italien, SC – Sizilien/Italien, SLO – Slowenien, AT – Österreich, HR – Kroatien, SRB – Serbien, BIH – Bosnien-Herzegowina, BG – Bulgarien, HUN –Ungarn, T – Thüringen
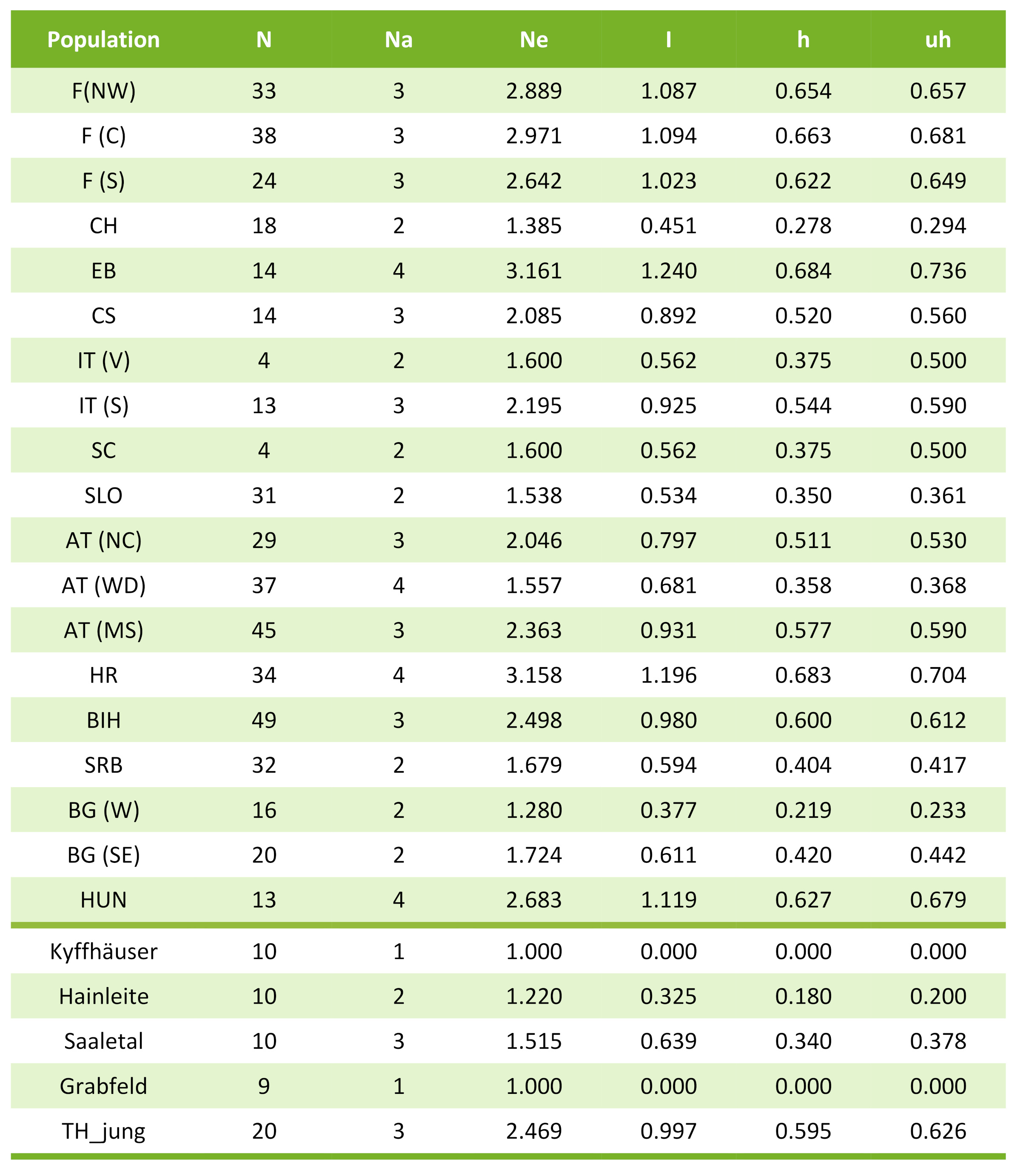
Während in allen untersuchten Populationen von Thüringer Altbäumen der Haplotyp 266 (blau) dominierte, stellte dieser in der Stichprobe der Jungspeierlinge nur den kleinsten Anteil. Insgesamt war die Haplotypendiversität in den Altbäumen (mit Ausnahme der Population "Saaletal") mit nur 1-2 Haplotypen sehr gering.
Im Gegensatz zu den Altspeierlingen zeigten die Jungspeierlinge eine Haplotypenzusammensetzung, die ungefähr einer Mischung aus dem mittel- bis osteuropäischen Durchschnitt entspricht. Der relativ große Anteil von Haplotyp 222 (rot) könnte auf einen eher osteuropäischen Ursprung hinweisen, allerdings ist die Schärfe des Markers bzw. auch die Stichprobengröße nicht groß genug um eine solche Vermutung zweifelsfrei abzusichern. Insgesamt ist auch die genetische Vielfalt (uh) bei den Jungspeierlingen deutlich höher als bei den Altspeierlingen (0.150 vs. 0.626, vgl. auch Tab. 1).
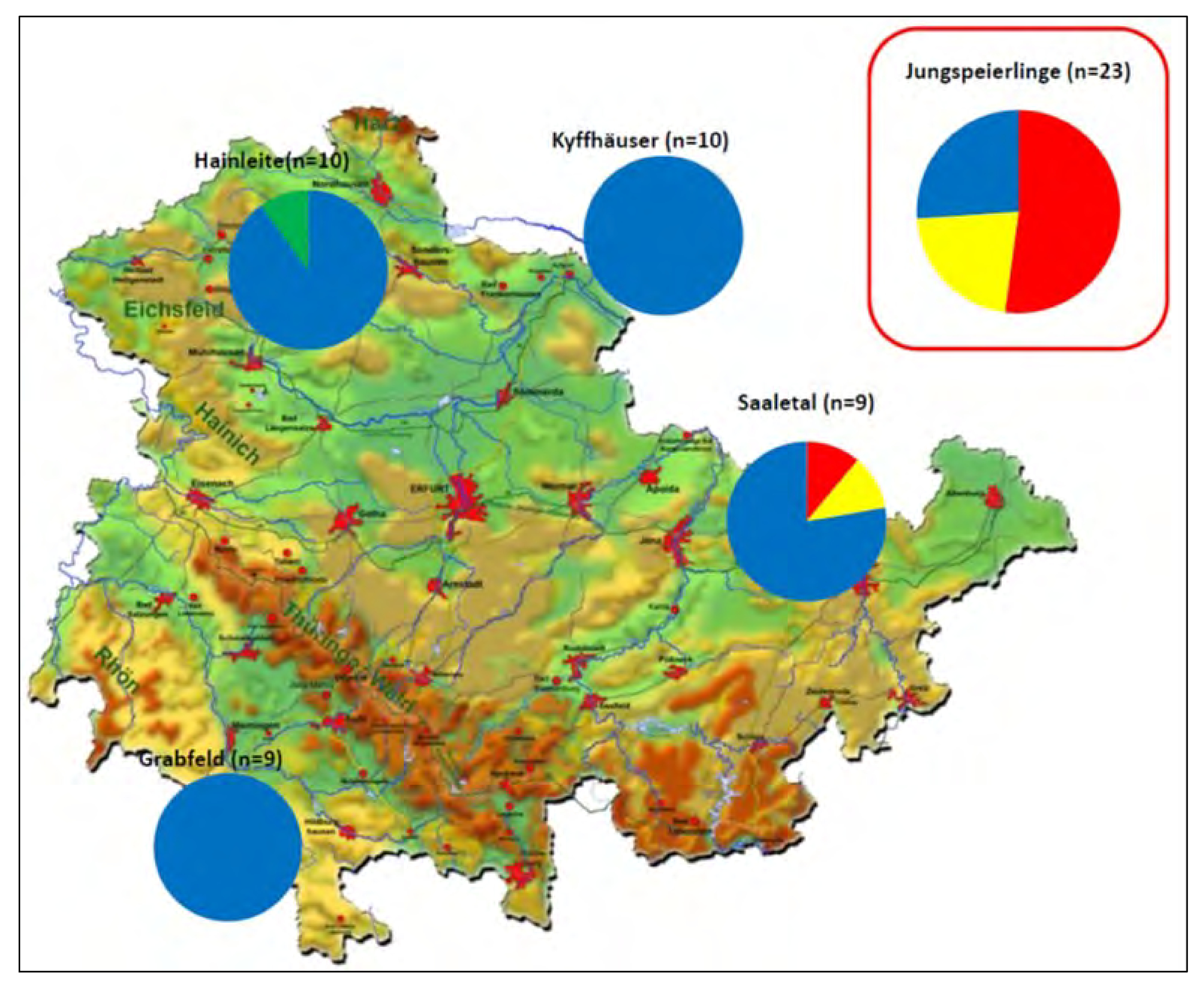
Abb. 1: Chloroplasten-Haplotypenverteilung innerhalb der Stichprobe aus Thüringen
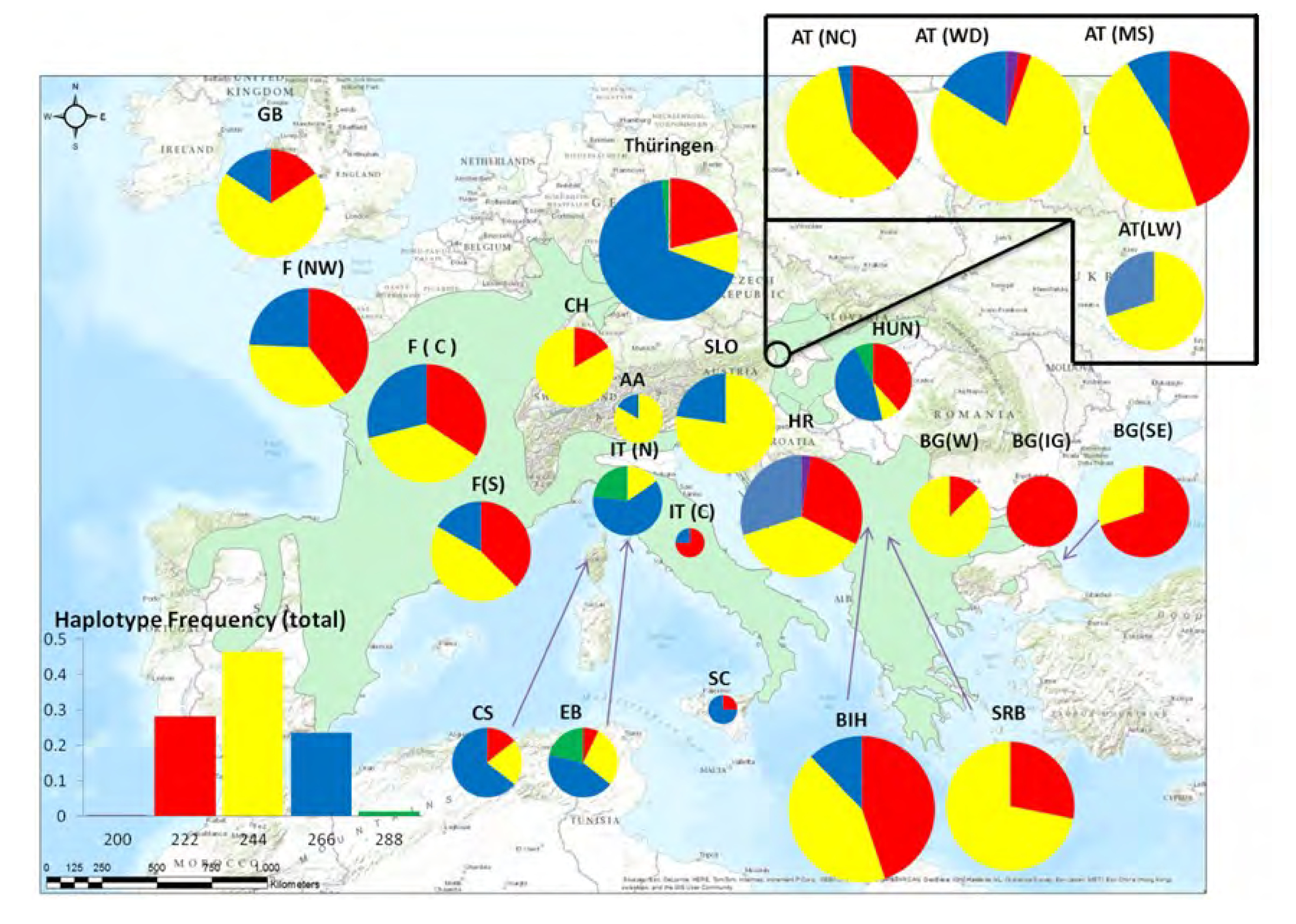
Abb. 2: Haplotypenverteilung der Thüringer Speierlinge (GER) im Vergleich zur gesamteuropäischen Stichprobe
Genetische Variation an nuklearen Mikrosatelliten
Die Ergebnisse der genetischen Variation untersucht anhand der nuklearen Mikrosatelliten-Marker sind in Tab. 2 dargestellt.
Tab. 2: Populationsdiversitätsindices (+/- Standardabweichung) aus den nuklearen Mikrosatellitendaten (7 Loci). Ne, Anzahl effektive Allele; Ho, beobachtete Heterozygotie; uHe, um die Stichprobengröße korrigierte erwartete Heterozygotie; F, Fixationsindex
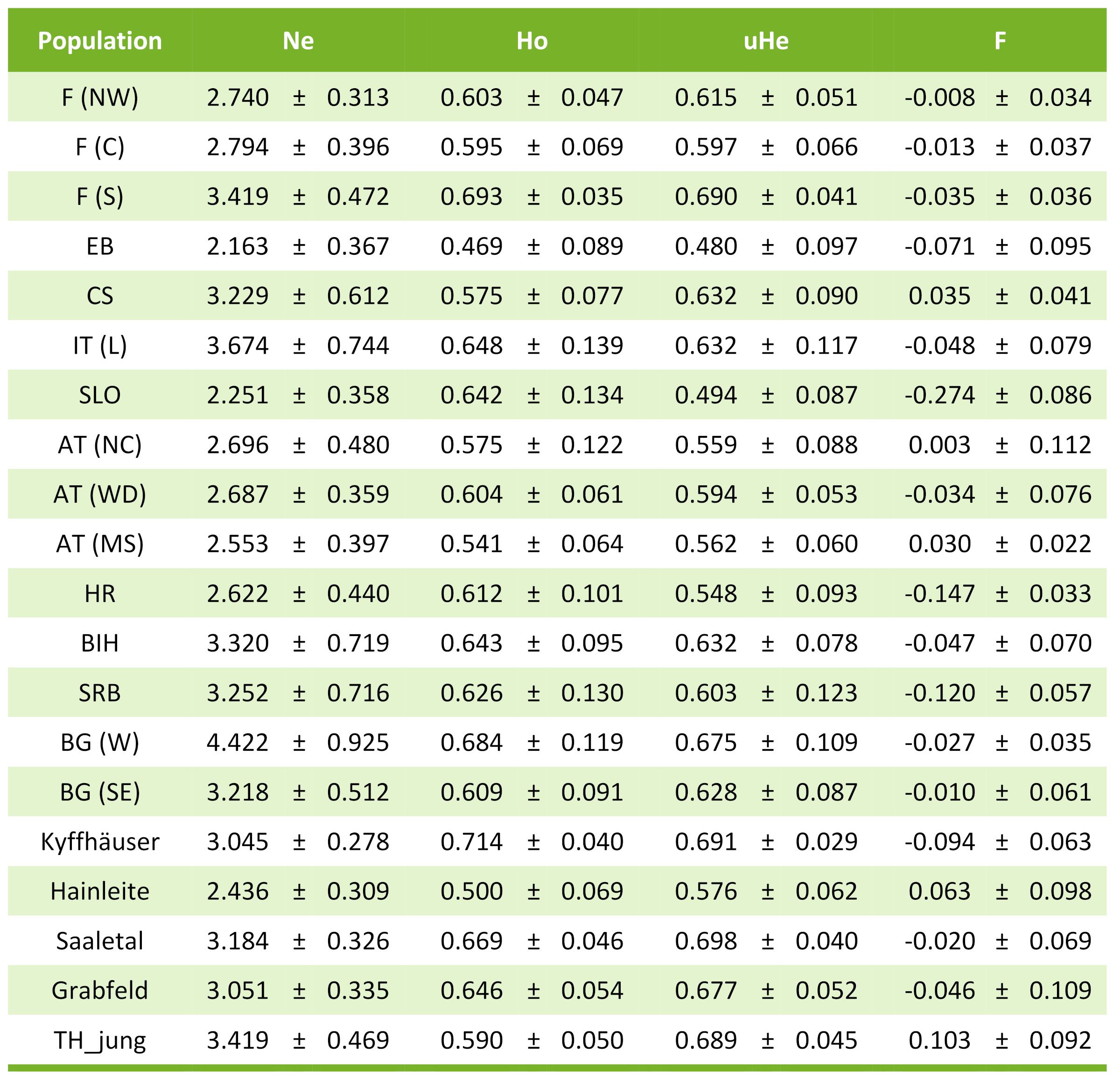
Die Untersuchung an 7 nuklearen Mikrosatelliten ergab, dass alle Thüringer Populationen eine für die Art relativ hohe genetische Vielfalt aufwiesen (im Durchschnitt erwartete Heterozygotie 0.688 bei Altspeierlingen bzw. 0.689 bei den Jungspeierlingen); lediglich die Population Hainleite zeigte hier einen niedrigeren Wert. Letztere Population weist auch einen positiven Fixationsindex auf, was ebenfalls ein Hinweis auf genetische Verarmung ist (Kreuzungen zwischen verwandten Individuen). Bei den anderen untersuchten Thüringer Teilpopulationen ist der Inzuchtwert (F) ausgeglichen, was auf das auskreuzende Paarungssystem hinweist (Inzucht wird dadurch weitestgehend verhindert).
Diskussion
Der in den Thüringer Altspeierlingen vorherrschende Chloroplasten-Haplotyp ist besonders in Westeuropa und im Mittelmeergebiet häufiger verbreitet; dies stimmt auch mit dem bekannten Verbreitungsgebiet der Art in Mitteleuropa überein, sodass eine Besiedlung (womöglich durch den Menschen auch begünstigt) über Frankreich nach Thüringen am wahrscheinlichsten erscheint. Die zusätzliche (spätere) Verbringung von Vermehrungsgut aus weiteren Regionen nach Thüringen ist ebenso möglich und wahrscheinlich, dafür würde auch das Vorkommen des Haplotypen 291 (grün) hindeuten, der sonst fast nur in Italien gefunden wurde. Eine Besiedlung aus dem Osten ist aufgrund des geringen (und wohl nicht natürlichen) Vorkommens der Art in Tschechien eher unwahrscheinlich.
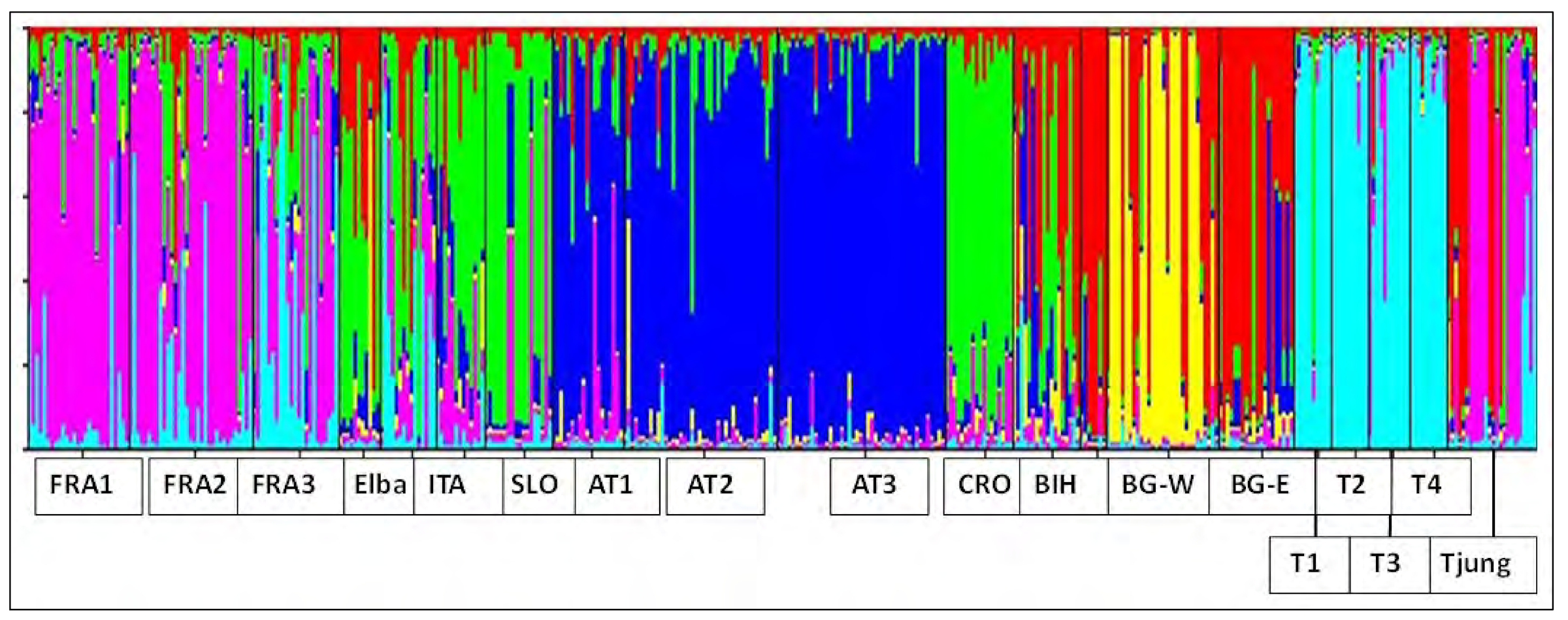
Abb. 3: Ergebnisse der auf individuellen Daten basierenden Zuweisung zu genetischen Gruppen mittels der Software STRUCTURE (PRITCHARD et al. 2000). Jede vertikale farbige Linie stellt die Wahrscheinlichkeit der Zuordnung zu einem bestimmten genetischen Cluster für ein Individuum dar, d.h. je signifikanter die Zuordnung eines Individuums zu einem Cluster umso „einfarbiger“ ist die jeweilige Linie. FRA – Frankreich, ITA – Italien, SLO – Slowenien, AT – Österreich, CRO – Kroatien, BIH – Bosnien-Herzegowina, BG – Bulgarien, T – Thüringen
Interessant ist jedoch die relativ starke Differenzierung der Thüringer Altspeierlinge, im Vergleich zu anderen Populationen aus Europa (GEORGE et al. 2015), sowie auch im Verhältnis zu den gepflanzten Jungspeierlingen. Besonders deutlich wird dies in der auf individuellen Daten basierenden Zuweisung zu genetischen Gruppen mittels der Software STRUCTURE (vgl. Abb. 3): hier bilden die Thüringer Altspeierlinge im Verhältnis zu anderen in Europa gesammelten Populationen eine sehr einheitliche Gruppe. Am ehesten gibt es eine Beziehung zu französischen und insbesondere südfranzösischen Populationen, hier sind auch die genetischen Distanzmaße am geringsten (durchschnittlich 0.25 Nei’s korrigierte Distanz). Dies stimmt auch mit der oben skizzierten Einwanderungsgeschichte der Art nach Deutschland und Thüringen und der bekannten aktuellen Verbreitung in Europa überein.
Insgesamt deuten die vorliegenden Ergebnisse darauf hin, dass die alten Speierlinge in Thüringen längere Zeit von anderen Vorkommen isoliert waren, andererseits sich aber auch (mehr oder weniger) natürlich vermehren konnten, da die Populationsstruktur der Altspeierlinge nur sehr wenige Abweichungen vom natürlich zu erwartenden Muster zeigten (nur geringe Abweichungen vom Hardy-Weinberg-Gleichgewicht). Nur wenige der Altbäume stammen aus anderen Regionen, etwa Frankreich und dem mediterranen Raum (Tab. 3).
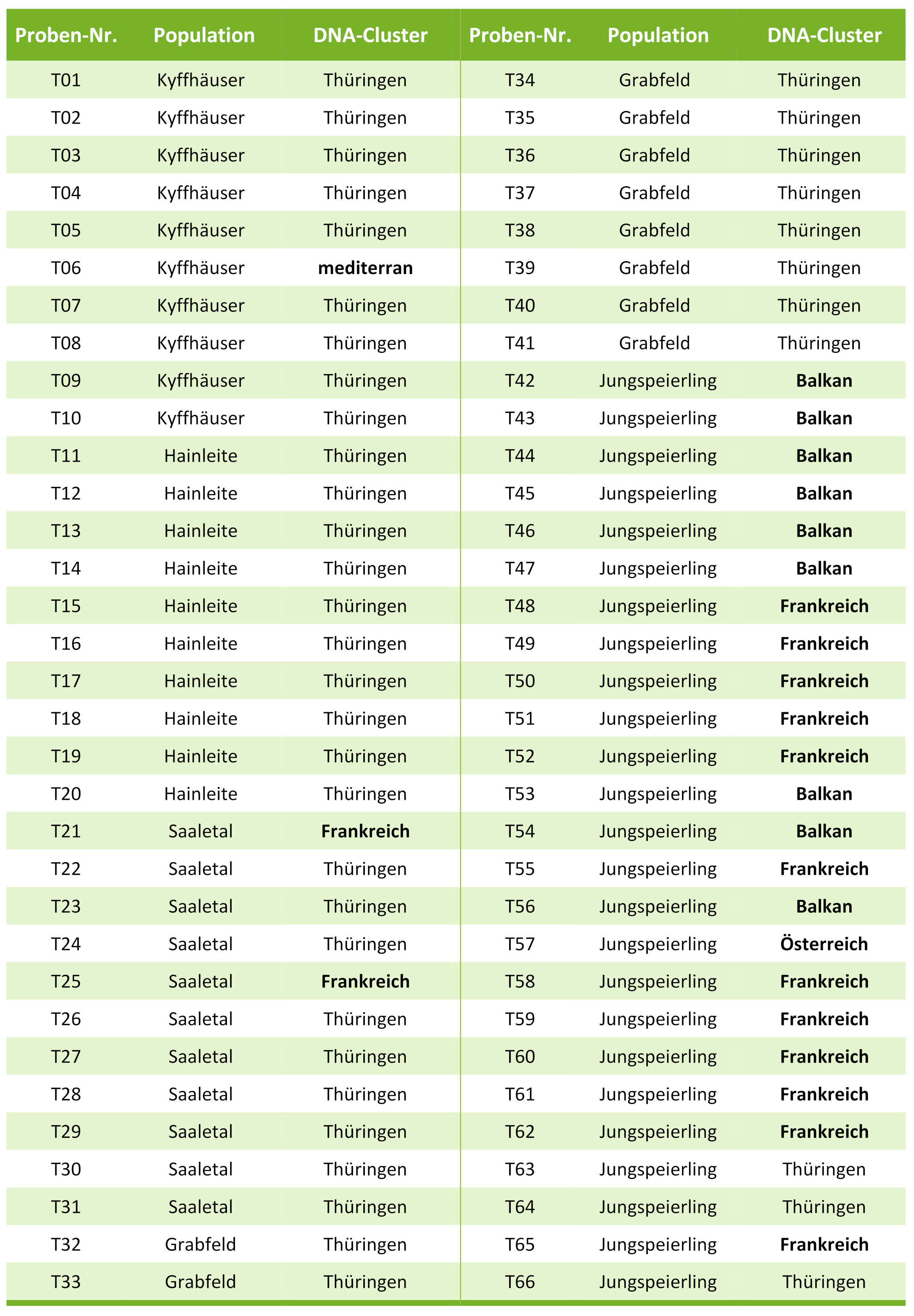
Tab. 3: Zuordnung der Thüringer Proben zu einem genetischen Cluster, basierend auf dem Ergebnis der STRUCTURE-Analyse der nuklearen Mikrosatelliten-Markerdaten
Im Gegensatz zu den Altspeierlingen weisen die Jungspeierlinge ein sehr heterogenes Muster in der STRUCTURE-Analyse auf (Abb. 3, Tab. 3). Dies ist vermutlich darauf zurückzuführen, dass hier Vermehrungsgut aus mehreren Teilen Europas verwendet worden ist, insbesondere dürften hier Herkünfte aus Frankreich bzw. aus Osteuropa (ehemaliges Jugoslawien) verwendet worden sein. Dies deckt sich auch gut mit den Ergebnissen des Chloroplasten-Markers. Drei der Jungspeierlinge (siehe Tab. 3) dürften aber aus der Thüringer Population stammen, offenbar wurde hier schon darauf geachtet den heimischen Genpool zu nutzen und zu erhalten.
Der Speierling ist ein gutes Beispiel dafür, wie wichtig es für die Erhaltung einer Art ist, Grundkenntnisse über ihre genetische Vielfalt zu haben bzw. gewisse Grundvoraussetzungen bei der Beerntung von Saatgut einzuhalten. In den letzten Jahrzehnten sind vermutlich tausende Speierlinge in Deutschland gepflanzt worden, aber oft wurde dabei auf deren Herkunft nur wenig geachtet. Meist wird günstig Saat- oder Pflanzgut aus anderen Ländern oder von großen Baumschulbetrieben eingekauft und dieses Material stammt dann aus Populationen, die große genetische Unterschiede zur heimischen Population aufweisen und womöglich auch weniger gut an die lokalen Bedingungen angepasst sind. Dazu kommt noch, dass es für die seltenen Baumarten in den meisten Ländern keine gesetzlichen Vorgaben gibt, die die Sammlung von Saatgut regulieren; d.h. Saatgut kann hier überall gesammelt und undeklariert verkauft werden. Im schlimmsten Fall kann ein Einzelbaum beerntet werden und dieses Saatgut wird dann für die Produktion tausender Jungpflanzen verwendet. Oft weist daher Handelsware nur eine sehr geringe genetische Vielfalt auf.
Genfluss in geringem Ausmaß dagegen hilft den fragmentierten Reliktpopulationen wiederum genetische Vielfalt aufzubauen, da sie ansonsten langfristig durch Inzuchterscheinungen vom Aussterben bedroht sind. Andererseits kann eine lokale autochthone Population in kürzester Zeit ausgelöscht werden, wenn in kleinen überalterten Vorkommen anstatt heimischer Jungpflanzen Material aus weit entfernten Regionen gepflanzt wird. Eine solche „Überschwemmung“ des heimischen Genpools (LEFÈVRE 2004) sollte auf jeden Fall vermieden werden, da dies dem Erhaltungsgedanken völlig zuwiderläuft. Eine derartige Entwicklung ist aber bei der gegenständlichen Untersuchung der Verjüngung deutlich erkennbar.
Abschließend kann gesagt werden, dass die gewonnen Ergebnisse eine überaus interessante Bereicherung unseres Wissensstandes über die Art darstellen. Die Thüringer Population zeigt Hinweise auf natürliche Entstehung und könnte so die nördliche Verbreitungsgrenze der Ausbreitung der Art nach Nordosten hin zeigen. Eine genetische Einengung an einer solchen Front, die theoretisch wahrscheinlich wäre, ist aber nicht zu erkennen, sondern sogar eher das Gegenteil davon. Die Art ist daher in Thüringen absolut erhaltungs- und schützenswert, die Verwendung von einheimischem Vermehrungsgut sollte auf jeden Fall gefördert werden.


