
Großflächige Schadereignisse wie Sturmwurf, Borkenkäferkalamitäten oder Schneebruch werfen immer wieder die Frage nach der Zweckmäßigkeit von kostenintensiven Wiederaufforstungsmaßnahmen auf. Meist stehen den Forstbetrieben nicht die nötigten finanziellen und personellen Mittel zur Verfügung, um in kurzer Zeit die Schadholzaufarbeitung und Aufforstung zu gewährleisten. Da der plötzliche Bestockungsverlust eine Entkopplung der Nährstoffkreisläufe, eine vermehrte Kohlenstoffdioxid- und Nährstofffreisetzung oder die Änderung der Wasserregime zur Folge hat (Wohlgemuth et al. 1995, Burschel & Huss 1997, Röhrig et al. 2006), gilt es die Flächen schnellstmöglich wieder zu bewalden. Die Pflicht zur Wiederaufforstung stark verlichteter Waldbestände ist daher auch im §11 Abs.1 BWaldG verankert und wird in den Landeswaldgesetzen nochmals konkretisiert (z.B. in Thüringen §23 ThürWaldG).
Im Zuge der Wiederbewaldung bieten Vorwaldstrukturen von Pionierbaumarten aus Naturverjüngung eine kostensparende Alternative zur Aufforstung durch Kunstverjüngung, da sie sich bei ausreichender Samenbaumanzahl im Zuge von Sukzessionsprozessen natürlicherweise einstellen. Pionierbaumarten vermögen aufgrund ihrer Arteigenschaften die Folgen der Freiflächenwirkung bereits nach kurzer Zeit zu mindern. Langfristige ökologische Defizite, die unter anderem ihre Ursache im Anbau einschichtiger Fichtenreinbestände haben, können durch Pionierbaumarten kompensiert werden (siehe Perala & Alm 1990, Schiechtl 1992, Prien 1995, Schmidt 1998, Hacker 1999, Schmidt 1999, Raspé et al. 2000, Argus 2006, Leder et al. 2007, Zerbe 2009, Hynynen et al. 2010). Außerdem ermöglicht die Einbindung von Pionierbaumarten in das Bestandesgefüge die Begründung naturnaher, standortsgerechter und stabiler Mischwälder.
Die Abschätzung des Besiedlungserfolgs durch Pionierbaumarten ist jedoch aufgrund begrenzter verjüngungsökologischer Kenntnisse zu maximalen Ausbreitungsdistanzen, optimalen Keimungsbedingungen oder standörtlichen Ansprüchen der Verjüngungspflanzen mit großen Unsicherheiten verbunden (u.a. Richter & Leder 1990, Lässig et al. 1995, Schmidt-Schütz & Huss 1998). Um zukünftig den Entscheidungsprozess zur Abwägung der Dringlichkeit von Maßnahmen auf Schadflächen - auch im Hinblick auf die Kalamitätsvorsorge in Fichtenwäldern - sicherer zu gestalten, wurden folgende Forschungsschwerpunkte in der vorliegenden Studie auf Kyrill-Sturmwurfflächen in den Hoch- und Kammlagen des Thüringer Waldes berücksichtigt:
- anemochore Samenausbreitung von Birke und Salweide sowie endozoochore Samenausbreitung durch Vogelarten auf Sturmwurfflächen;
- Potenzial der Bodensamenbanken von Pionierbaumarten auf Sturmwurfflächen und in fichtenbestockten Waldflächen;
- Verjüngungsfortschritt von Pionierbaumarten auf Sturmwurfflächen;
- Stichprobenhafte Erhebung der aktuellen Samenbaumvorkommen von Pionierbaumarten in den Hoch- und Kammlagen des Thüringer Waldes.
Untersuchungsgebiet und Versuchsdesign
Das Untersuchungsgebiet entspricht der ausgewiesenen Fläche des Modellprojektes "Waldumbau in den mittleren, Hoch- und Kammlagen des Thüringer Waldes", die zwischen Oberhof, Zella-Mehlis, Schmiedefeld und Ilmenau gelegen ist (Abb. 1). Das Gebiet des Modellprojektes umfasst eine Fläche von ca. 8.000 ha und wird dem Wuchsgebiet „Thüringer Gebirge“ und dem Wuchsbezirk „Mittlerer Thüringer Wald“ zugeordnet (Burse et al. 1997). Der Wuchsbezirk stellt sich vorrangig als Kammgebirge mit einem hohen Hanglagenanteil und häufig fehlenden Plateaulagen dar (Burse et al. 1997, Waesch 2003, Gauer & Aldinger 2005).

Abb. 1: Lage der Untersuchungsflächen im Gebiet des Modellprojektes "Waldumbau in den mittleren, Hoch- und Kammlagen des Thüringer Waldes"
Die Aufnahmen wurden auf sieben standörtlich vergleichbaren, ehemals fichtenbestockten Kyrill-Sturmwurfflächen (Januar 2007) mit Ausdehnungen der damaligen Blößen von 4,0 - 12,7 ha in den Hoch- und Kammlagen (750 - 900 m ü. NN) durchgeführt (Abb. 1 und Tab. 1). In einem 500 m breiten Streifen um die Sturmwurfflächen wurden alle Birken-, Salweiden- und Ebereschensamenbäume erfasst. Unter Berücksichtigung der jeweiligen Flächenausdehnung und -form wurden kreuz- und sternförmig angeordnete Linientransekte angelegt. Die Aufstellung von Samen- und Kotfallen (Abb. 2), die Entnahmen der Bodenproben und die Verjüngungsaufnahmen erfolgten entlang dieser Transekte stets in Abständen von 20 m. Die Kotfallen wurden dabei jeweils auf freien Flächenbereichen und unter einem, im unmittelbaren Umkreis vorhandenen Strukturelement platziert (siehe Tiebel et al. 2017).

Abb. 2: Netztrichterfalle für Birkensamenfänge (links), Kotfalle für Ebereschensamen (Mitte) und Klebfallen für Weidensamenfänge (rechts) (Fotos K. Tiebel)
Zudem wurden Bodenproben in Fichtenbeständen mit vereinzelt eingemischten Ebereschen- und Birkensamenbäumen entnommen. Abschließend wurde in dem Untersuchungsgebiet auf einer Teilfläche von 600 ha eine Samenbauminventur durchgeführt. Die Aufnahmen erfolgten entsprechend eines 200 x 200 m Rasters. Zusätzlich konnten alle Samenbäume der Pionierbaumarten entlang befahrbarer Waldwege erfasst werden (Stähr et al. 2006, Hosius et al. 2012). Alle Untersuchungen fanden im Zeitraum von 2014 - 2016 statt.
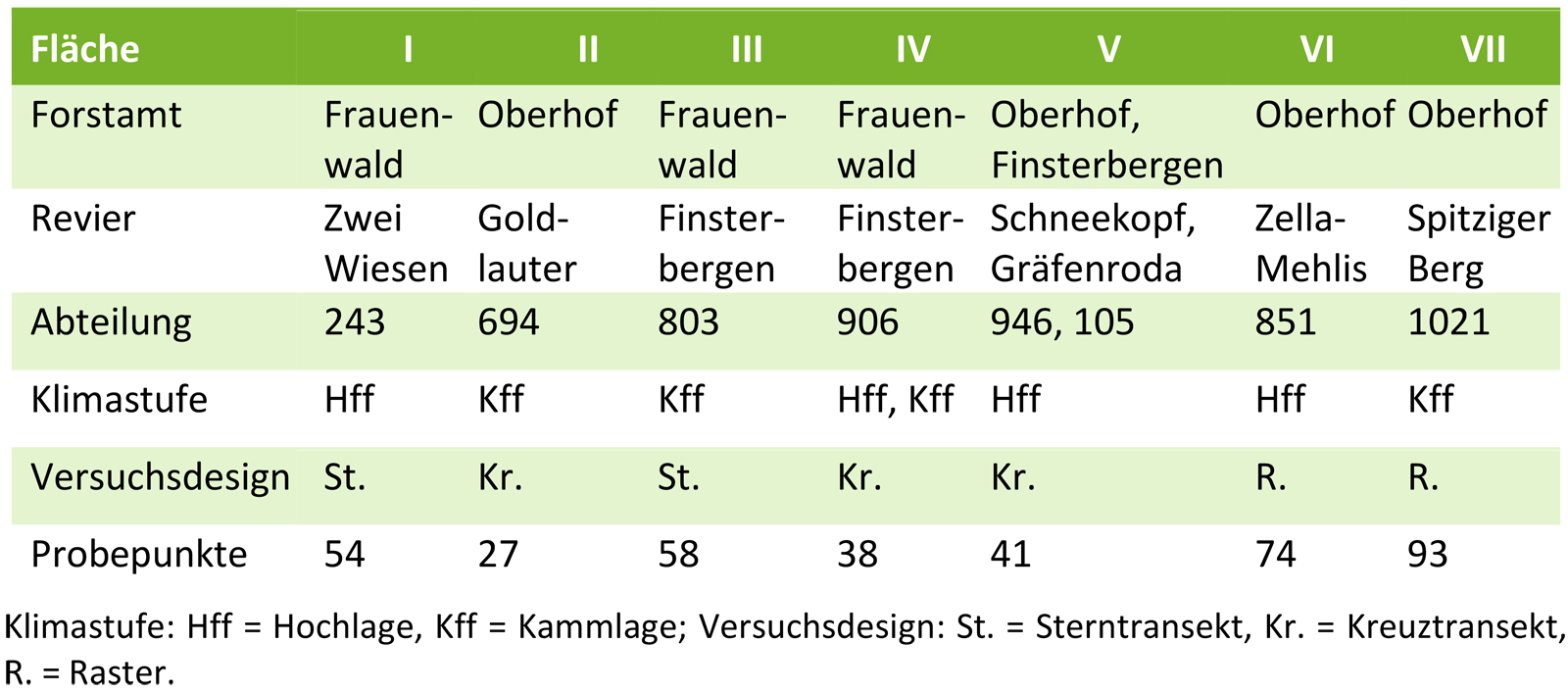
Tab. 1: Übersicht über die Untersuchungsflächen und das Versuchsdesign.
Samenausbreitung von Birke und Salweide
Ergebnisse
Der Samenflug von Birke und Salweide wurde auf den gleichen Sturmwurfflächen in zwei aufeinanderfolgenden Jahren, 2015 und 2016, untersucht. Die erfassten Samendichten fielen für beide Pionierbaumarten im Jahr 2015 signifikant geringer aus (U-Test: p<0,003). Für Birke wurden auf den Sturmwurfflächen mittlere Samendichten von 8,3 - 92,5 n m-2 (2015) nachgewiesen. Im Jahr 2016 erreichte die mittlere Samendichte Werte zwischen 46 - 445 n m-2 (Abb. 3 links). Die Samendichte betrug damit das Fünffache im Vergleich zum Vorjahr. Die maximal erfassten Dichten beliefen sich auf 2.053 (2015) und 9.557 n m-2 (2016).
Bei Salweide betrugen die Fangzahlen 2015 zwischen 0,6 - 1,8 n je Falle und im Jahr 2016 zwischen 1,1 - 2,1 n je Falle, womit der Unterschied nicht so deutlich wie bei Birke ausfiel (Abb. 3 rechts). Die höchste Samendichte von 156 n je Falle (2016) wurde direkt unter einem Samenbaum der Salweiden gemessen. Auf den samenbaumfreien Sturmwurfflächen erreichten die maximalen Samendichten 5 n je Falle(2015) und 9 n je Falle (2016).
Kenntnisse über die mittleren Ausbreitungsdistanzen und Richtungseffekte liefert für Birke die inverse Modellierung zur Samenausbreitung (siehe van Putten et al. 2012). Dabei ergaben sich für den richtungsunabhängigen Fall mittlere Ausbreitungsdistanzen (MDD) von 86 - 380 m. Für Salweide wurden Ausbreitungsdistanzen bis 870 m und darüber hinaus nachgewiesen. Ein mathematischer Richtungseffekt (Windfahne) konnte für beide Pionierbaumarten nicht belegt werden.
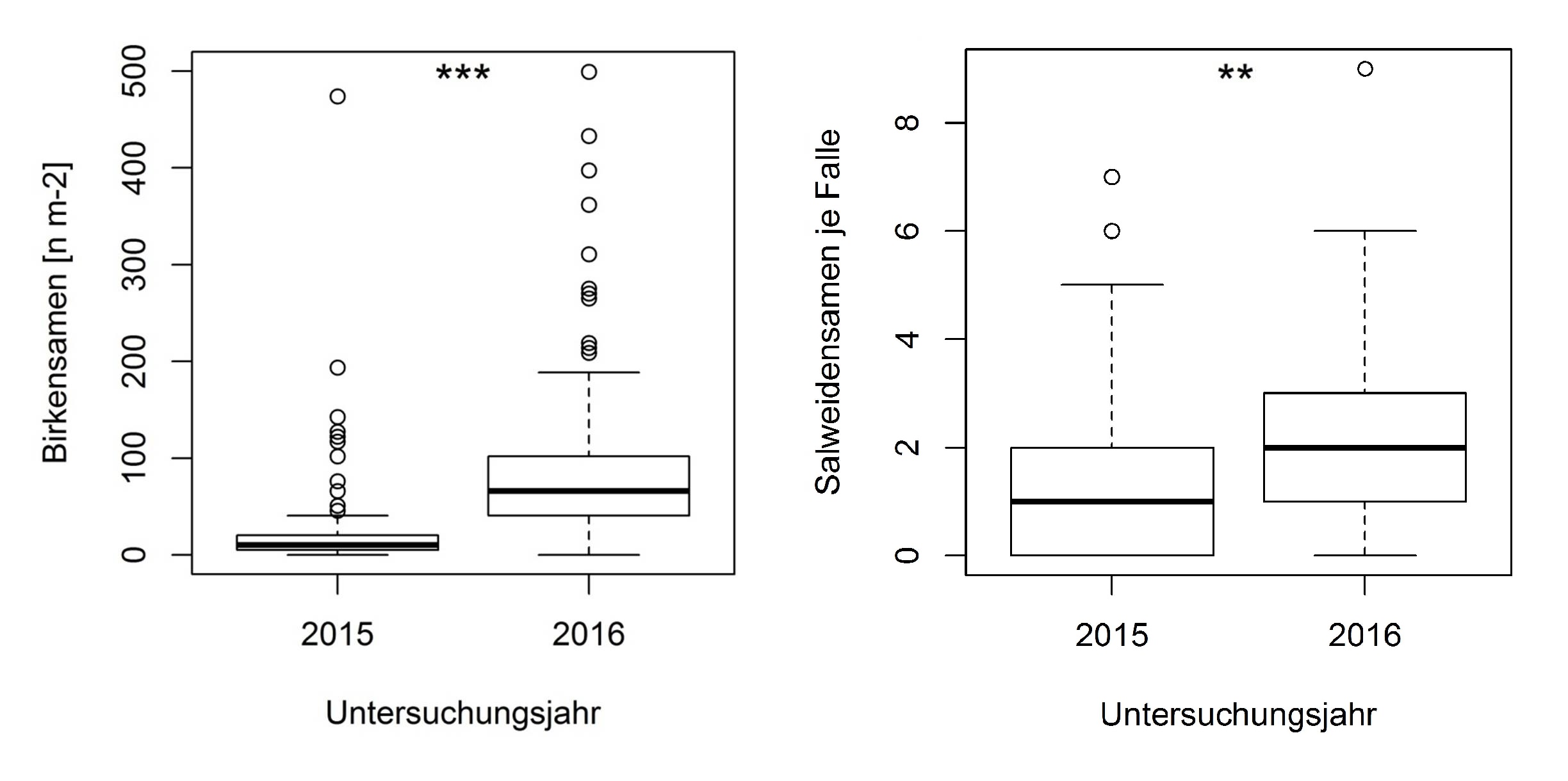
Abb. 3: Birkensamendichten [n m-²] und Salweidensamendichten je Falle in den Jahren 2015 und 2016 (H-Test: p<0,01**)
Schussfolgerungen für die Forstpraxis
Die Qualität und Quantität der jährlichen Fruktifikation sowie die zeitlichen und räumlichen Ausbreitungsmuster windverbreiteter Baumarten wie Birke werden durch vielfältige Faktoren und Rahmenbedingungen beeinflusst. Dimension, Kronenausdehnung (bzw. Konkurrenzdruck und Pflegerückstände) und individuelle Konstitution eines Samenbaumes sind entscheidende Einflussgrößen bezüglich der absolut produzierbaren Samenmengen eines Baumes (Buschel & Huss 1997). Zudem können von Jahr zu Jahr deutliche Unterschiede in den produzierten Samenmengen auftreten, ähnlich einem Mastjahr. Diese unterschiedlichen Samenmengen sind u.a. durch variierende Witterungsbedingungen begründet (Sarvas 1952, Bastide & van Vredenburch 1970, Atkinson 1992). Aus den Untersuchungsergebnissen geht hervor, dass mit einer Besiedlung von Störungsflächen mit waldbaulich relevanten Verjüngungsdichten von Birke vorrangig in ausgewählten Jahren mit größeren Mengen produzierter Samen („Mastjahren“) zu rechnen ist.
Die räumlichen Ausbreitungsmuster der abgelagerten Birkensamen auf den Sturmwurfflächen zeigen, dass die höchsten Samendichten jeweils in Samenbaumnähe zu finden sind. Ebenso unterliegt die Samenausbreitung dem Einfluss des Reliefs, aber nicht der vorherrschenden Windrichtung. So betrug die hangabwärts gerichtete mittlere Ausbreitung 380 m, wohingegen die Birkensamen hangaufwärts mit 97 m eine deutlich geringere mittlere Distanz zurücklegten. Folglich kommt auch der Position und der räumlichen Verteilung der Samenbäume um eine Störungsfläche eine große Bedeutung zu.
Die Untersuchungen zur Samenausbreitung vonSalweide haben gezeigt, dass (i) Richtungseffekte keine Rolle bei der Samenausbreitung spielen, (ii) die Position des Samenbaums im Gelände keinen Einfluss hat (=Reliefunabhängigkeit) und (iii) die Samenbaumanzahl auf die abgelagerte Samendichte ab einer Entfernung von > 50 m vernachlässigbar ist. Im Zuge der enormen Fernausbreitung lösen sich zeitliche und räumliche Ablagerungsmuster auf. Die Ergebnisse lassen den Schluss zu, dass von einer samenquellenunabhängigen Samenablagerung zwischen 10 - 20 n m-² während des jährlichen Reproduktionszeitraums ausgegangen werden kann. Mit Blick auf die natürliche Wiederbewaldung von Störungsflächen durch Salweide sind die erfassten Samendichten somit ausreichend. Limitierende Faktoren bei der Wiederbewaldung sind nicht die Flugweiten der Salweiden, sondern (i) Wildverbiss, (ii) fehlende oder unzureichend vorhandene Mikrostandorte und (iii) klimatisch-standörtlich ungünstige Bedingung während der Keimung. Salweidensamen verlieren bereits nach wenigen Tagen bis Wochen unter natürlichen Bedingungen ihre Keimfähigkeit (HECKER 1998, SCHÜTT et al. 2011).
Eine räumliche Optimierung von Samenbäumen zur prophylaktischen Stärkung der Resilienz in kalamitätsgefährdeten Fichtenwäldern muss lediglich für die Birke vorgenommen werden. Dabei gilt es möglichst viele, vitale Samenbäume verteilt auf der Fläche vorzuhalten. Durch die weiten Ausbreitungsdistanzen und Omnipräsens der Samen trifft dies nicht für Salweide zu. Allerdings müssen für beide (alle) Pionierbaumarten waldbauliche Maßnahmen zur Erhaltung, Vitalisierung und Förderung neuer Samenbäume in die Behandlungskonzepte der Bestände integriert werden.
Endozoochore Samenausbreitung durch Vogelarten
Ergebnisse
Die Untersuchungen zur endozoochoren Samenausbreitung durch Vogelarten auf Sturmwurfflächen unter Freiflächenbedingungen und Strukturelementen fanden im Jahr 2015 statt. Aufgrund des Versuchsaufbaus waren nicht an jedem Aufnahmepunkt vergleichbare Strukturelemente zur Beprobung verfügbar (Abb. 4). Die vorgefundene Vielfalt der Strukturelemente bot jedoch die Möglichkeit, Präferenzen der Vogelarten bei der Wahl von Rast- und Sitzgelegenheiten zu benennen.
Abbildung 4 belegt den häufigeren Aufenthalt frugivorer (=Früchte fressende) Vögel auf abgestorbenen Baumteilen. Anhand der Kotdichte am Boden konnte gezeigt werden, dass bevorzugt Totäste (20 n m-²) aufgesucht wurden. Unter umgeklappten Wurzeltellern erreichte die mittlere Kothaufendichte 4,6 n m-². Im Vergleich zu den Stubben (0,8 n m-²) mit einer maximalen Höhe von einem Meter wurden unter Hochstubben (3,9 n m-²) deutlich höhere Kothaufendichten nachgewiesen. Im direkten Umfeld oder unter bereits etablierter Verjüngung von Birke und Eberesche fand sich kein Vogelkot. Zwischen den Fallenpunkten unter Fichtenverjüngung auf den Sturmwurfflächen (1,1 n m-²) und unter Fichten der Wuchsklasse Baumholz in den angrenzenden Beständen (1,8 n m-²) ließen sich keine eindeutigen Unterschiede hinsichtlich der Kothaufendichte dokumentieren. Unter den künstlich eingebrachten Sitzgelegenheiten (Einzelschutz und Zaun) war kein Vogelkot vorhanden. Insgesamt konnte eine signifikant höhere Dichte an Vogelkot (2,7 n m-²) auf Kotfallen unter Strukturelementen im Vergleich zu Kotfallen auf den Freiflächen ohne Strukturelemente nachgewiesen werden (U-Test: p=0,000). An allen Aufnahmepunkten ohne Strukturelemente erreichte die mittlere Kothaufendichte lediglich 0,4 n m-².
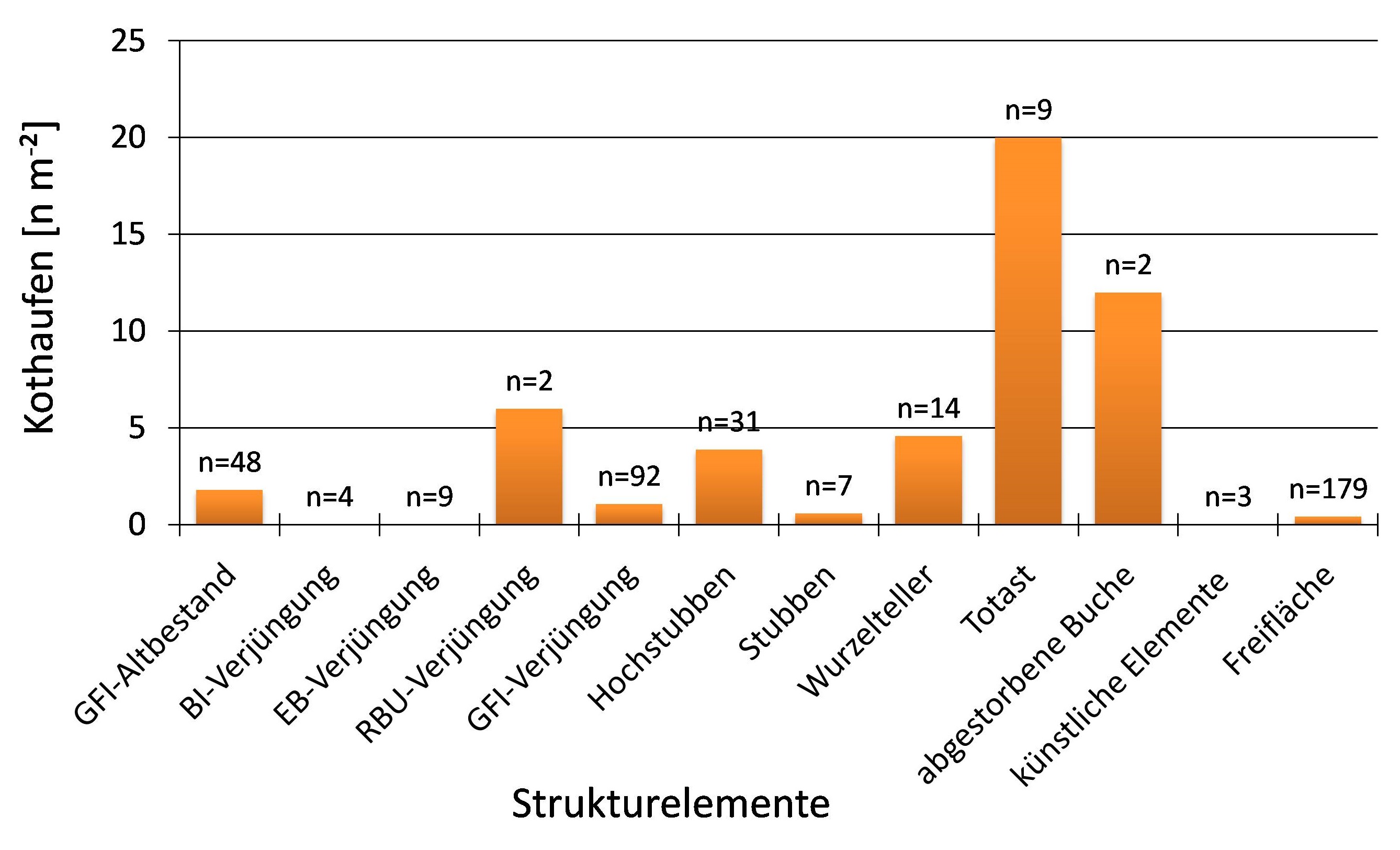
Abb. 4: Mittlere Vogelkotdichten [n m-²] unter den beprobten Strukturelementen und unter Freiflächenbedingungen
Schussfolgerungen für die Forstpraxis
Das Vorhandensein von Strukturelementen ist eine notwendige Voraussetzung für den endozoochoren Sameneintrag durch Vögel auf Freiflächen. Für eine gezielte natürliche Wiederbewaldung von Sturmwurfflächen mit einer ausreichenden Verjüngungsdichte an endozoochor verbreiteten Baumarten reicht die nachgewiesene Vogelkotdichte unter „realen“ Freiflächenbedingen ohne vorhandene Rast- und Sitzgelegenheiten nicht aus (u.a. McDonnell & Stiles 1983, Jordano & Schupp 2000).
Wie die Studie belegen konnte, bevorzugen frugivore Vogelarten Rast- und Sitzgelegenheiten zum Absetzen von Kot auf Freiflächen. Sie bieten oftmals Sichtschutz vor Feinden und zugleich ein weites Sichtfeld (McDonnell & Stiles 1983, Kollmann 2000, Żywiec & Ledwoń 2008). Die vorliegende Untersuchung weist dabei auf eine Präferenz der Vögel für stehende, stärker dimensionierte und mindestens einen Meter hohe Strukturelemente hin. Schwach dimensionierte Verjüngungspflanzen der Birke, Eberesche oder Fichte wurden auf den untersuchten Sturmwurfflächen weitgehend von den Vogelarten gemieden bzw. nicht zum Absetzen von Kot aufgesucht (siehe Tiebel et al. 2017). Künstlich eingebrachte Sitzgelegenheiten stellen nur unter bestimmten Voraussetzungen eine Alternative dar. Maßgeblich sind vor allem vertikale Strukturen mit horizontalen Abzweigungen und ausreichenden Sitzhöhen von mindestens anderthalb Meter (McDonnell & Stiles 1983, McDonnell 1986).
Für den praktischen Umgang mit Strukturelementen auf Störungsflächen lässt sich schlussfolgern, dass eine komplette Beräumung der „Schlüsselstrukturen“ vermieden werden sollte (Abb. 5). Eine Reduktion der strukturellen Diversität ist weder aus ökonomischer noch aus ökologischer Sicht zu rechtfertigen. Vor allem durch das Belassen von: (i) liegendem und stehendem Totholz mit stärker dimensionierten Ästen, (ii) einer ausreichenden Anzahl an beasteten Hochstubben und (iii) geworfenen Wurzeltellern können bestmögliche Voraussetzungen für den Sameneintrag auf Sturmwurfflächen geschaffen werden. Bereits etablierte Verjüngungspflanzen endozoochor verbreiteter Baumarten auf Störungsflächen sollten als potentielle Samenbäume möglichst erhalten werden. Fruchttragende Baumarten, wie die Eberesche, weisen den Vorteil einer frühen und reichlichen Fruktifikation im Freistand oder bei Freistellung auf, sodass diese Bäume zeitnah zur räumlichen Vernetzung der Samenquellen beitragen können (Gockel 2016).

Abb. 5: Etablierte Ebereschenverjüngung um den Stammfuß einer Altfichte (Foto K. Tiebel)
Bodensamenbank
Ergebnisse
Die Untersuchungen zum Bodensamenbankpotential von Pionierbaumarten wurden für Eberesche und Birke durchgeführt (Abb. 6). Für Eberesche erbrachten die Untersuchungen keine Ergebnisse. Unabhängig von den beprobten Flächen waren keine Samen dieser Baumart im Boden nachweisbar.

Abb. 6: Schematische Darstellung der Probenaufbereitung des gestochenen Bodens bis zur fertigen Keimprobe (Fotos K. Tiebel)
Birkensamen waren dagegen für alle untersuchten Standorte nachweisbar. In den Fichtenbeständen mit einzelbaumweise eingemischten Birken fanden sich durchschnittlich 48 n m-² keimfähige Samen, wovon 20 n m-² auf den Mineralboden und 28 n m-² auf die Auflagehorizonte entfielen (Tab. 2). In reinen Fichtenbeständen ohne Einzelbäume der Birke belief sich die Birkensamendichte auf lediglich 3 n m-². Diese befanden sich ausnahmslos in den Auflagehorizonten. In den Mineralbodenproben der Sturmwurfflächen wurden nur auf einer Fläche in unmittelbarere Nähe zum Samenbaum keimfähige Birkensamen (8 n m-²) gefunden. Auf diesen Flächen wurden jedoch nicht die Auflagenhorizonte untersucht. Unabhängig von den beprobten Standorten, nahmen die Birkensamendichten in den Proben mit zunehmender Bodentiefe signifikant ab (H-Test: p<0,001).
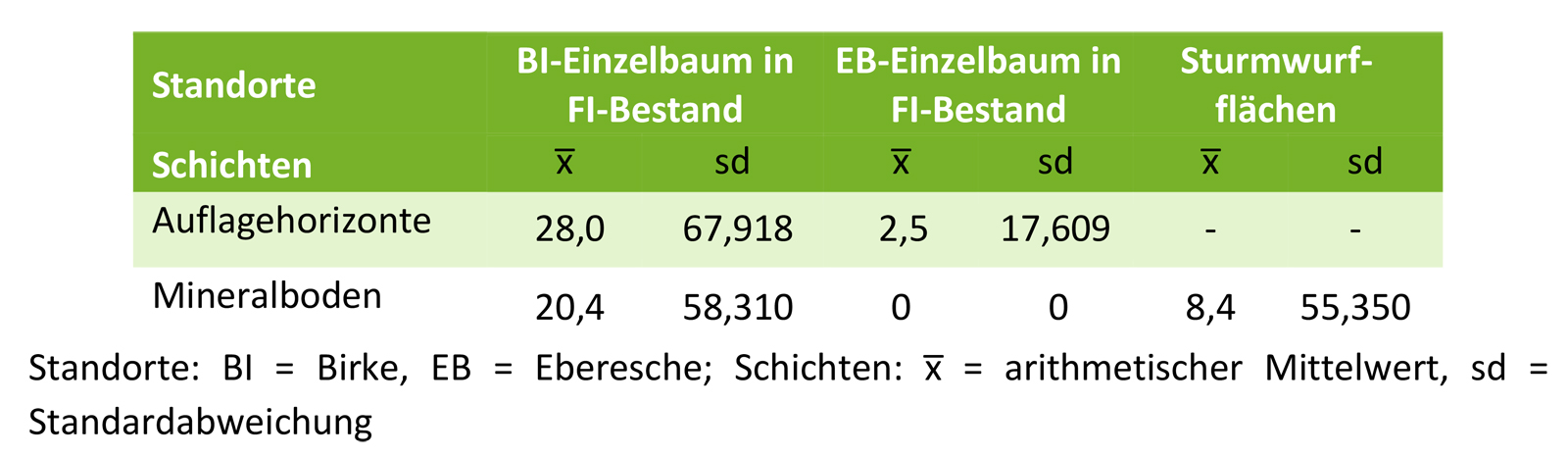
Tab. 2: Birkensamendichten [n m-²] in den untersuchten Proben der Bodensamenbank, getrennt nach den Untersuchungsflächen und Bodenschichten.
![Balkendiagramm zur Darstellung der relativen Häufigkeit verbissener Individuen [%] in den einzelnen Verbissklassen (=Verbissprozente an den Individuen), getrennt nach den Baumartengruppen](/files/_processed_/6/2/csm_Tiebel_et_al_Abb7_519314458d.webp)
Abb. 7: Darstellung der relativen Häufigkeit verbissener Individuen [%] in den einzelnen Verbissklassen (=Verbissprozente an den Individuen), getrennt nach den Baumartengruppen
Schussfolgerungen für die Forstpraxis
Bodensamenbanken setzen sich unabhängig vom betrachteten Ökosystem hauptsächlich aus früh- und mittelsukzessionalen Arten zusammen, sogenannten lichtbedürftigen Arten oder Störungszeigern. Diese Arten sind zum Aufbau einer persistenten Bodensamenbank befähigt (Donelan & Thompson 1980, Bossuty et al. 2002). Die Untersuchungen auf den Sturmwurfflächen zeigen, dass die ehemalige Bodensamenbank mit dem Störungsereignis im Januar 2007 aktiviert wurde. Aufgrund des fehlenden Nachweises von Birkensamen im Mineralboden müssen ehemals im Boden befindliche Samen nach dem Sturmereignis gekeimt sein oder zum Zeitpunkt der Untersuchung nicht mehr keimfähig gewesen sein. Der Nachweis von Birkensamen in der transienten Bodensamenbank (Auflagehorizonte) deutet dagegen auf ein neu im Aufbau befindliches Bodensamenbankreservoir hin. Wahrscheinlich werden erst mit zunehmender Bestandesentwicklung keimfähige Birkensamen in tiefere Bodenschichten gelangen.
Keimfähige Samen von Eberesche wurden in den Bodenproben der verschiedenen Untersuchungsflächen nicht gefunden, obgleich diese Baumart die einzige Pionierbaumart ist, deren Samen nachweislich 3 - 5 Jahre im Boden überliegen kann (Hill 1979, Leder 1992, Erlbeck 1998). Birkensamen waren dagegen auf allen Flächen nachweisbar, obwohl viele Autoren sich gegen die Bodensamenbankfähigkeit dieser Baumart aussprechen (Hill & Stevens 1981, Amezaga & Onaindia 1997, Buckley et al. 1997, Ebrecht & Schmidt 2008, Heinrichs 2010). Komulainen et al. (1994) schreibt jedoch, dass in Mastjahren von Birke große Mengen abgelagerter Birkensamen im Boden akkumuliert werden können, wenn die Keimbedingungen unvorteilhaft sind. Dies erklärt die hohen Samendichten in der transienten Bodensamenbank. Allerdings zeigt die signifikante Abnahme keimfähiger Samen mit zunehmender Bodentiefe auch eine begrenzte Verweilzeit der Birkensamen im Boden. In verschiedenen Studien wurde die Birke infolge der divergierenden Ergebnisse unterschiedlichen Bodensamenbanktypen zugeordnet (siehe Tiebel et al. 2018). Die Angaben zur Erhaltung der Keimfähigkeit im Boden reichen dabei von weniger als einem Jahr bis zu 4 - 5 Jahren (Bakker et al. 1996, Thompson et al. 1997, Bekker et al. 2000). Um eine kontinuierliche Verjüngung zu gewährleisten, muss die Bodensamenbank daher so oft wie möglich aus umliegenden Samenbäumen gespeist werden.
Verjüngungsetablierung
Ergebnisse
Die erfassten Verjüngungsdichten auf den Kyrill-Sturmwurfflächen der Hoch- und Kammlagen betrugen für Birke 0,06 - 0,27 n m-² und Eberesche 0,06 - 0,88 n m-². Demgegenüber belief sich die Verjüngungszahl von Salweide und Zitterpappel lediglich auf 0 - 0,2 n m-² und 0 - 0,12 n m-².
Die erfassten Individuen unterlagen zudem einem art- und flächenspezifisch unterschiedlich starken Verbissdruck durch Wild. Die baumartenspezifischen mittleren Verbissraten lagen bei 19 %, 62 %, 65 % und 85 % für Birke, Eberesche, Pappel und Weide. Detailliertere Angaben zum Verbissdruck sind aus der Abbildung 7 zu entnehmen.
Schussfolgerungen für die Forstpraxis
Im Rahmen der Vegetationsaufnahmen wurden 8 Jahre nach Sturmwurf Verjüngungsdichten von Pionierbaumarten zwischen 4.200 - 11.700 Individuen pro ha erfasst. Bei der Gegenüberstellung mit vergleichbaren Studien ergab sich eine ausreichende Verjüngungsdichte im Sinne eines Vorwaldcharakters (siehe Falinska 1998, 1999, Keidel et al. 2008, Wohlgemuth & Kramer 2015). Laut den Maßgaben von ThüringenForst galt eine Sturmwurffläche nach Sturm Kyrill bei einer Verjüngungsdichte von vitalen und unbeschädigten Individuen mit > 2.000 Laubbäume pro ha (= 0,2 n m-²) oder > 1.000 Nadelbäume pro ha (= 0,1 n m-²) als ausreichend wiederbewaldet (TMLNU 2009). Zum Zeitpunkt der Vegetationsaufnahmen erfüllten alle Sturmwurfflächen die quantitativen Vorgaben, obgleich die Verjüngungspflanzen oft in geklumpter Form und nicht homogen über die Fläche verteilt waren (Abb. 8). Allerdings muss infolge des extrem hohen Verbissdrucks bei fortschreitender Flächenentwicklung mit einer weiteren Reduktion der Verjüngungsdichten gerechnet werden, die sich aufgrund höherer Mortalitätsraten nach zu starker Schädigung der Pflanzen einstellt. Nach Aussagen von Keidel et al. (2008) sterben Ebereschen bei einem Terminaltriebverbiss von mehr als 80 % mit großer Wahrscheinlichkeit ab. Mit Blick auf den bereits vorhandenen dichten Begleitwuchs krautiger Pflanzen, die ein Aufwachsen neuer Pionierbaumarten erschweren, kann eine bisher als ausreichend verjüngte Fläche dadurch wieder in die Kategorie „ungenügend gesicherte Verjüngung“ fallen.
Samenbauminventur
Ergebnisse
Die Stichprobeninventur auf 600 ha zur Erhebung der Samenbaumvorräte von Pionierbaumarten im Thüringer Wald ergab eine deutliche Dominanz der Eberesche. Insgesamt konnten in dem Gebiet 200 Ebereschen-Samenbäume lokalisiert werden, wovon 131 Individuen unmittelbar am Wegrand stockten. Dies entsprach einem Mittelwert von 10 n km-1 Weg. Dagegen wurden 8 Birken- und 16 Salweiden-Samenbäume gefunden, wovon lediglich zwei Salweiden und eine Birke an Aufnahmepunkten innerhalb der Bestände erfasst wurden.

Abb. 8: Mit Birke erfolgreich wiederbewaldete Kyrill-Sturmwurffläche (Foto K. Tiebel)
Des Weiteren traten die Samenbäume aller untersuchten Pionierbaumarten sowohl am Weg, als auch in den Waldflächen in aggregierter Form auf. Die höchsten Samenbaumdichten innerhalb der ausgewählten Stichprobenfläche wies das Revier Schneekopf im Bereich Schmücke und Schneekopf auf (Abb. 9). In diesem Revier waren auch alle detektierten Samenbäume der Salweiden und Birken angesiedelt. Die Eberesche wurde hingegen auch vereinzelt in den benachbarten Revieren Goldlauter, Zella-Mehlis und Zwei Wiesen nachgewiesen. Keine Samenbäume von Birke, Salweide und Eberesche fanden sich dagegen im Revier Finsterberg.
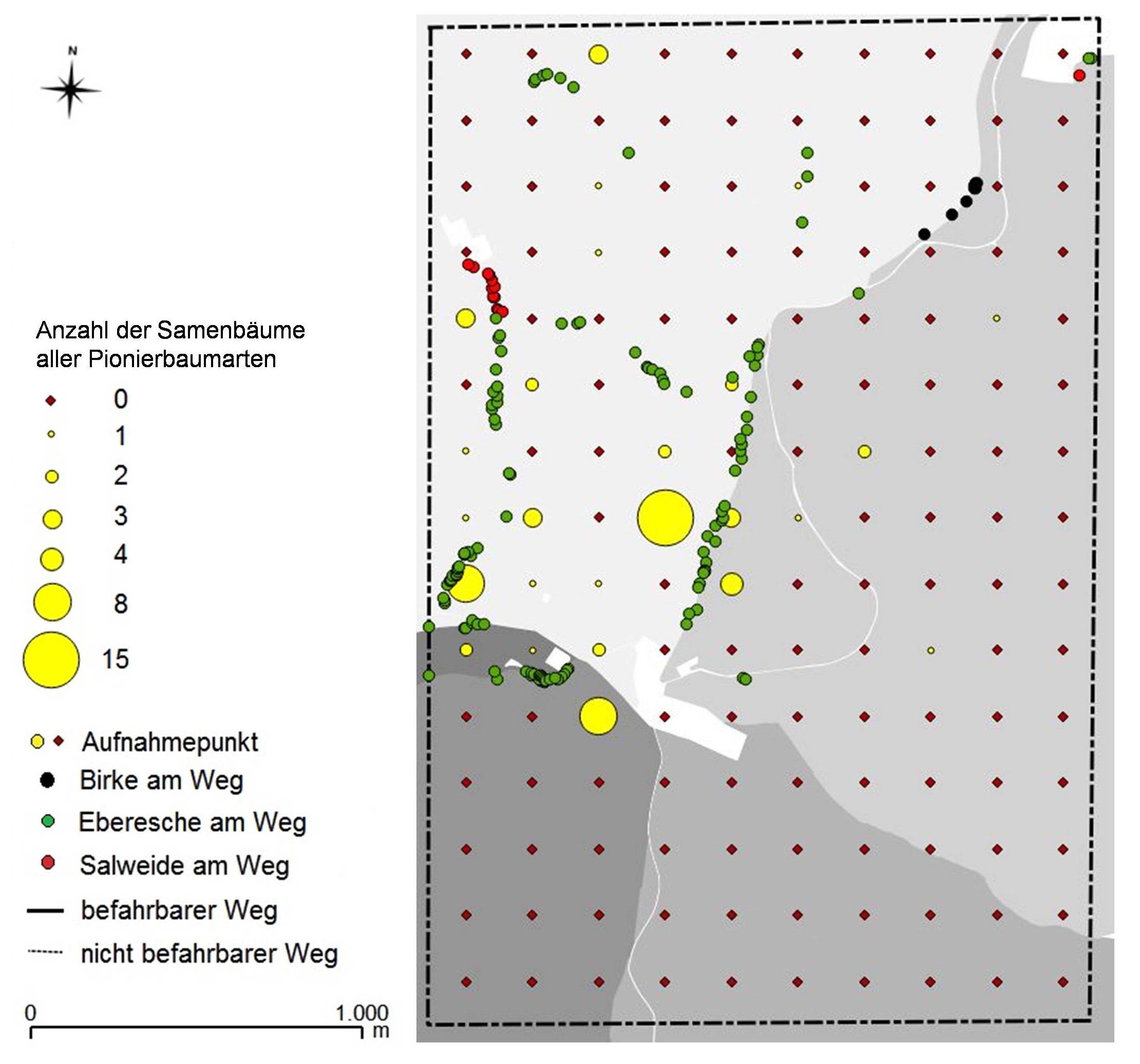
Abb. 9: Kartografische Darstellung der Samenbaumpotenziale an den Aufnahmepunkten (200 x 200 m) und befahrbaren Waldwegen auf der 600 ha großen Teilfläche im Untersuchungsgebiet
Schussfolgerungen für die Forstpraxis
Die Ergebnisse der Samenbauminventur haben ein unzureichendes Samenbaumpotenzial im Falle neuer Schadkatastrophen für einen großen Bereich des untersuchten Waldgebietes offengelegt. Besonders häufig fehlte es an Birken und Salweiden. Mit Ausnahme der ehemals aktiv geförderten Individuen an den Wegrändern waren diese Baumarten kaum in den Beständen zu finden. Aus diesem Grund ist die Schaffung von Lückeninitialen in den sonst geschlossenen Nadelwaldbeständen zur Begründung neuer Samenbaumpotentiale unabdingbar, sowie eine aktive Förderung vorhandener Individuen empfehlenswert (Atkinson 1992, Perala & Alm 1990).
Auch die Eberesche ist mit Ausnahme des Revieres Schneekopf in den Waldflächen stark unterrepräsentiert. Die große Dichte von Ebereschen-Samenbäumen im Bereich Schneekopf und Schmücke ist der lokalen Flächengeschichte als sowjetisches Sperrgebiet geschuldet. Pionierbaumarten brauchten nicht aktiv aus Forstkulturen entnommen werden, wie es für die Wirtschaftswälder zu dieser Zeit vorgegeben war (Röhrig & Gusson 1990, Leder 1992). Dieser flächenspezifische Sonderfall zeigt jedoch, dass Ebereschen, wenn sie im Bestand belassen werden, mit dem Hauptbestand aufwachsen und sich etablieren können. Allerdings gilt es, wie gleichermaßen für Birke und Salweide auch, die Baumarten aktiv zu fördern, indem Individuen oder Gruppen freigestellt werden. Denn alle Ebereschen-Samenbäume in den Beständen wiesen schlecht entwickelte Kronen, schlechte Qualitäten, geringe Stabilitäten, geringe Vitalitäten und mitunter erste Absterbeerscheinungen auf. Dies sind die Folgen einer fehlenden Pflege in der Vergangenheit. Dichtstand und mangelhafte Kronenentwicklung geht zudem häufig mit einer unzureichenden bis ausbleibenden Fruktifikation einher (Gockel 2016). Für das angestrebte Ziel der natürlichen Wiederbewaldung im Falle neuer Schadkatastrophen stellen die zweifelsfrei vorhandenen Ebereschen-Samenbäume demnach keine optimale Ausgangssituation dar.
Zur Schaffung ausreichender Samenbaumpotentiale von Pionierbaumarten in den fichtenwald-dominierten Flächen gilt es zukünftig (i) Verjüngungsinitiale zu schaffen, (ii) den Schutz der Verjüngung vor Wildverbiss zu gewährleisten und (iii) etablierte Individuen oder Gruppen turnusmäßig zu pflegen und damit die Kronenentwicklung sowie Einzelbaumstabilität zu fördern.