

Abb. 1. Edellaubbaumarten, hier ein Mischbestand aus Berg-Ahorn, Gemeiner Esche und Spitz-Ahorn, sind im Erzgebirge häufig durch eine artenreiche Epiphytenflora gekennzeichnet. Foto: Martin Baumann (SBS)
Einen wesentlichen Bestandteil der pflanzlichen Diversität von Waldökosystemen stellen epiphytische Moose dar. Dies ist dadurch bedingt, dass viele dieser Arten sehr substratspezifisch sind. Dabei zeigen epiphytische Moose eine Abhängigkeit von der Trägerbaumart. Als besonders epiphytenfreundlich gelten viele Laubbaumarten, z.B. Gemeine Esche, Ahorn- und Pappel-Arten. Dagegen sind Nadelbaumarten für die Besiedlung durch epiphytische Moose weniger gut geeignet.
Da epiphytische Moose ihre Wasser- und Nährstoffversorgung nahezu ausschließlich über das Niederschlagswasser decken, weisen sie eine besonders hohe Sensitivität gegenüber Schadstoff-, insbesondere gegenüber Schwefel (S) -Depositionen auf. Aufgrund der in Mitteleuropa generell hohen S-Depositionen im letzten Jahrhundert war diese Artengruppe stark rückläufig, vor allem im Umfeld der Industriezentren. Diese hohe Sensitivität machte man sich zunutze, um anhand der Artenzusammensetzung epiphytischer Moose auf die Luftgüte zu schließen. Mit dem Rückgang der S-Depositionen ist eine Wiederausbreitung epiphytischer Moosarten zu verzeichnen.
Ausgangslage in Sachsen
In der zweiten Hälfte des 20. Jahrhunderts zählt das Erzgebirge zu den Regionen mit den höchsten S-Depositionen in Mitteleuropa. Als Folge dieser extremen Luftverschmutzung starben grosse Fichtenbestände komplett ab und es kam zu einer tiefgründigen Versauerung der Waldböden, so dass die Vorkommen zahlreicher epiphytischer Moosarten als erloschen galten. Durch technische Maßnahmen nach der politischen Wende erfolgte eine deutliche Verbesserung der Luftqualität.
Historisch bedingt ist im Erzgebirge der Anteil der Fichte sehr hoch. Bei aktuellen Erhebungen wurden beispielsweise in den Forstbezirken Adorf, Eibenstock und Neudorf Fichtenanteile im Oberstand von 85,2 bis 86,2 Prozent festgestellt. Die Anteile der Rotbuche als Hauptbaumart der dominierenden natürlichen Waldgesellschaft betrug dagegen nur 2,7 bis 4,7 Prozent. Mit dem Übergang zu einem ökologisch orientierten Waldbau ab etwa 1985 begann ein systematischer Umbau der homogenen Nadelbaum-Forsten in standortgerechte Mischbestände. Im Erzgebirge erfolgte das überwiegend durch einen Voranbau von Rot-Buche, Weiß-Tanne, Berg-Ahorn etc. in Fichtenbeständen.
Zwei Untersuchungsziele
Ziel der vorliegenden Untersuchung war es,
die aktuelle Artenzusammensetzung epiphytischer Moose in den Waldökosystemen des Erzgebirges nach einer Wiederbesiedlungsdauer von etwa 15 Jahren zu erfassen,
- die Unterschiede in der Artenzusammensetzung auf verschiedenen Trägerbaumarten zu analysierne und zu bewerten.
Die Ergebnisse sollen Rückschlüsse hinsichtlich der Artenzusammensetzung und Diversität epiphytischer Moose erlauben, die infolge des Umbaus homogener Fichten- in Mischbestände unter Beteiligung von standortgerechten Laubbaumarten erwartet werden kann.
Verwendete Methoden
Grundlage für die Untersuchung bildete das Versuchsflächensystem der Baumart Gemeine Fichte im Erzgebirge. Es wurden zwölf Probeflächen mit einer Größe von jeweils 0,5 bis 1,0 ha ausgewählt (Abb. 2). Anschließend wurden im Umfeld Horste bzw. Kleinbestände weiterer Baumarten gesucht und Probeflächen in möglichst vergleichbarer Flächengröße festgelegt. Dabei handelte es sich zum einen mit Rot-Buche und Berg-Ahorn (Abb. 1) um die Hauptbaumart und eine wesentliche Mischbaumart des Hainsimsen-Buchenwaldes, welcher im Erzgebirge bis in die Hochlagen die bestimmende natürliche Waldgesellschaft darstellt. Zum anderen erfolgten die Untersuchungen auf Gemeiner Eberesche, einer Mischbaumart im Reitgras-Fichtenwald, der dominierenden natürlichen Waldgesellschaft in den Kammlagen. Die ausgewählten Probeflächen decken mit einer Meereshöhe von 550 bis 940 m die Klimastufen der Mittleren und Höheren Berglagen sowie die Kammlagen ab.
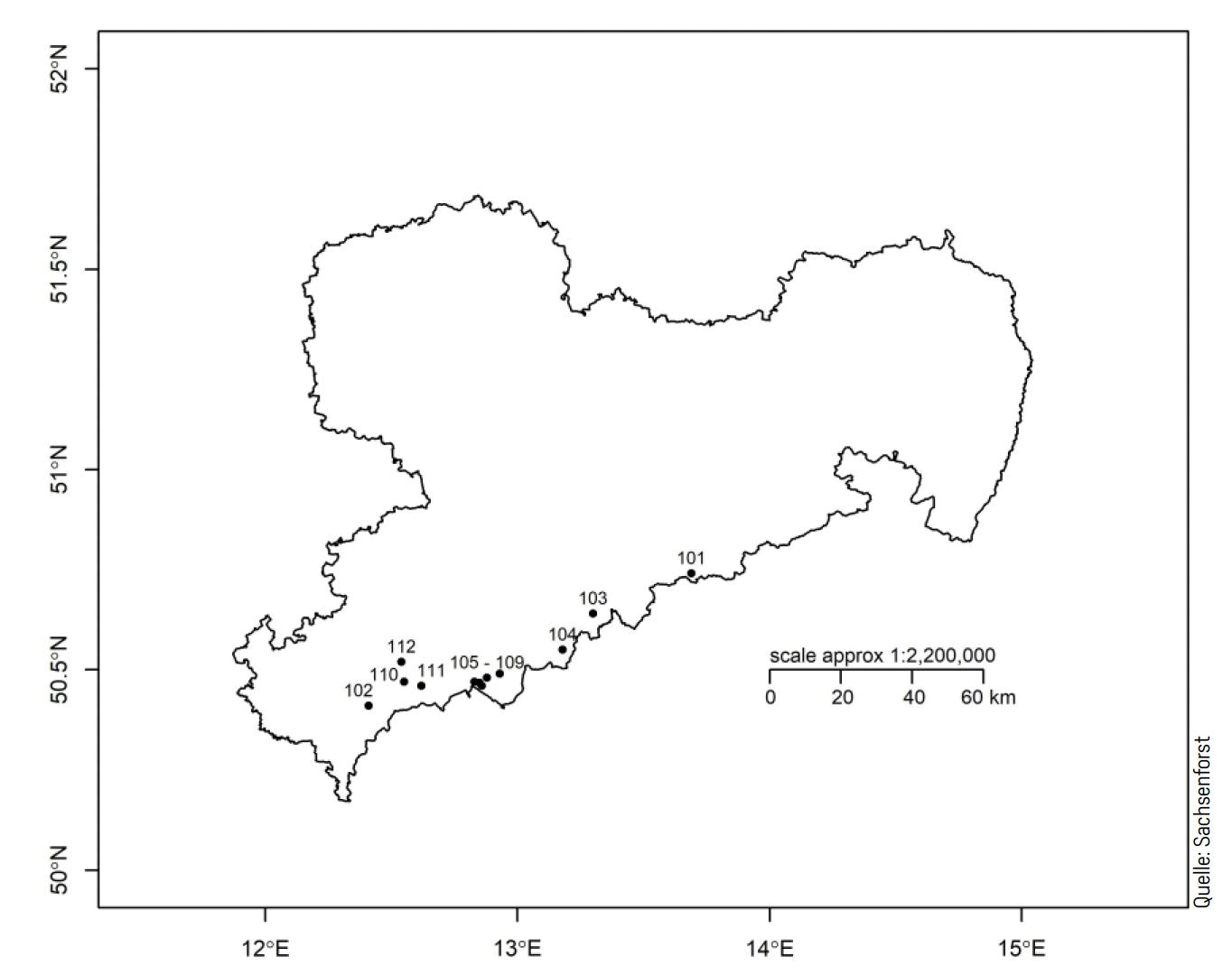
Abb. 2. Karte mit der Lage der untersuchten Fichten-Versuchsflächen im Territorium des Freistaates Sachsen. © SBS.
In jeder Probefläche wurden die vorkommenden epiphytischen Moose bis in Reichhöhe (Stammbereiche sowie Äste in einer Höhe zwischen ca. 0.5 und 2.0 m) erfasst.
Es wurden Parameter für die alpha- und gamma-Diversität epiphytischer Moose ermittelt. Während die alpha-Diversität die lokale Artenvielfalt (z.B. auf Ebene des Bestandes) darstellt, bezieht sich die gamma-Diversität auf die Landschaftsebene. Insofern wird als Maß für die alpha-Diversität die Artenzahl pro Probefläche betrachtet, als eine Schätzung für die gamma-Diversität die Artenzahl pro Baumart in allen Probeflächen (d.h. der regionale Artenpool). Diese Parameter sind zum einen für alle Arten, zum anderen für die Artengruppe der obligaten Epiphyten berechnet worden. Weiterhin erfolgte eine Analyse der gefährdeten Arten nach der aktuellen Roten Liste für Sachsen, inkl. aller Arten, die erst im 21. Jh. in Sachsen nachgewiesen worden sind. Außerdem wurde der prozentuale Anteil kurzlebiger Arten ermittelt.
Der zeitliche Verlauf der Wiederbesiedlung konnte durch einen Vergleich mit Daten aus dem Zeitraum zwischen 2005 und 2008 analysiert werden. Weitere Details finden sich im Originalartikel.
Untersuchungsergebnisse
Auf den Probeflächen wurden insgesamt 84 epiphytische Moosarten gefunden, wobei die Anzahl pro Probefläche zwischen 0 in einer Fichtenfläche und 37 in drei Ahorn- bzw. einer Buchenfläche schwankte. Die mittleren Artenzahlen fielen auf Eberesche, Buche und Ahorn signifikant höher aus als auf Fichte (Tab. 1). Dieselben Relationen ergaben sich, wenn nur die obligaten Epiphyten berücksichtigt wurden. Auf Fichte wiesen überwiegend substratunspezifische Arten sowie Arten mit Verbreitungsschwerpunkt auf Totholz bzw. Waldboden epiphytische Vorkommen auf. Auf den untersuchten Laubbaumarten dominierte dagegen die Artengruppe der obligaten Epiphyten.
Tab. 1. Diversitätsparameter auf Probeflächen der Baumarten Fichte, Eberesche, Buche und Ahorn, p-value: Ergebnis Kruskal-Wallis Test, ungleiche Buchstaben indizieren signifikante Unterschiede (Dunn`s Post-hoc-Test).
| Fichte | Eberesche | Buche | Ahorn | p-value | |
| lokaler Artenpool (alpha-Diversität) | 9.2a | 26.5b | 30.6c | 32.8c | <0.001 |
| (alle Arten) | ±7.02 | ±2.75 | ±3,78 | ±4.30 | |
| lokaler Artenpool (alpha-Diversität) | 3.3a | 16.6ab | 20.1c | 22.5c | <0.001 |
| (obligate Epiphyten) | ±4.41 | ±2.27 | ±4.60 | ±2.06 | |
| Anteil obligater Epiphyten in % | 35.9 | 62.6 | 65.7 | 68.6 | |
| regionaler Artenpool (gamma-Diversität) | 34 | 53 | 68 | 64 | |
| (alle Arten) | |||||
| regionaler Artenpool (obligate Epiphyten) | 17 | 31 | 39 | 39 |
Den höchsten Wert für den regionalen Artenpool wies die Buche auf. Für diese Baumart wurden auf allen Probenflächen 68 epiphytische Moose bzw. 39 obligate Epiphyten festgestellt. Ähnlich viele Arten besaßen die Ahornflächen, während die Fichte mit 34 Arten bzw. 17 obligaten Epiphyten nur halb so viele Arten im Vergleich zur Buche besaß. Zu den häufigen Arten zählt u.a. Hypnum cupressiforme mit 46 und Orthotrichum affine (Abb. 3) mit 42 Vorkommen. 19 Arten wiesen dagegen nur jeweils ein Vorkommen auf. Mit Ausnahme dieser seltenen Arten blieb keine epiphytische Moosart auf nur eine Baumart beschränkt. Nur sechs Arten liessen sich als Indikatorarten für eine Baumart identifizieren:
- Fichte: Plagiothecium curvifolium
- Eberesche: Rhytidiadelphus loreus, Ceratodon purpureus
- Ahorn: Orthotrichum anomalum, Ptychostomum moravicum, Cryphea heteromalla (s. Abb. 4)

Abb. 3. Zahlreiche epiphytische Arten gehören zur Gattung der Goldhaarmoose (Orthotrichum), die häufigste Art dieser Gattung im Erzgebirge ist aktuell Orthotrichum affine.Foto: S. Dittrich

Abb. 4. Das Einseitswendige Verstecktfruchtmoos (Cryphaea heteromalla) wurde erst 2009 zum ersten Mal in Sachsen gefunden. Seit zwei Jahrzehnten ist eine Ausbreitung der als submediterran-subatlantisch geltenden Art nach Osten und ins Binnenland zu verzeichnen. Foto: J. Nixdorf

Abb. 5. Das Gewöhnliche Igelhaubenmoos (Metzgeria furcata) gehört zu den thallosen Lebermoosen. Es wächst sowohl auf Rinde als auch kalk- bzw. basenreichem Gestein. Foto: M. Baumann (SBS)
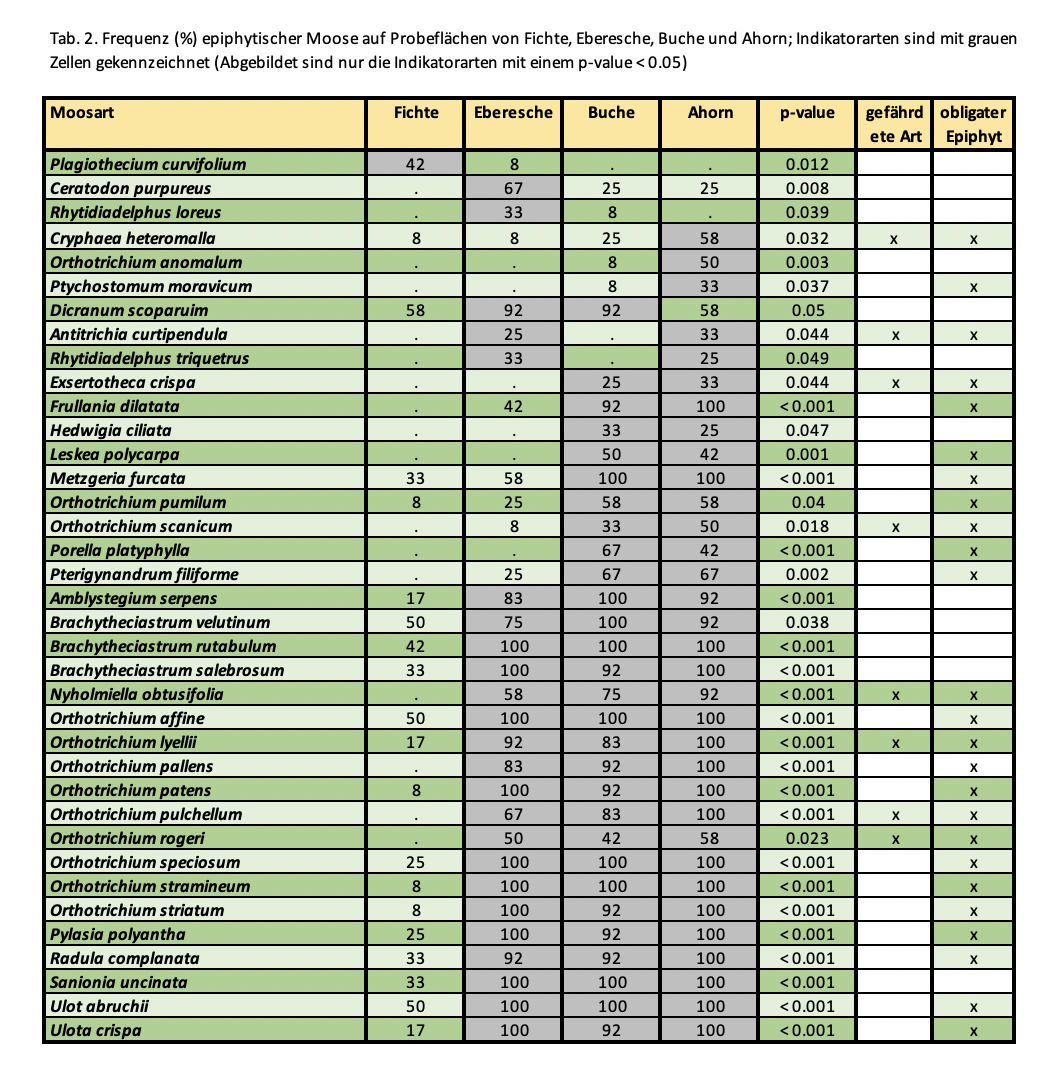
Dagegen zeigten 19 epiphytische Moose, davon 14 obligate Epiphyten, eine signifikante Präferenz für die drei untersuchten Laubbaumarten. Für Ahorn und Buche ließen sich weitere neun Indikatorarten identifizieren, darunter die drei Lebermoose Frullania dilatata, Porella platyphylla und Metzgeria furctata (Abb. 5). Insgesamt ist die Artenzusammensetzung epiphytischer Moose auf den untersuchten Laubbaumarten sehr ähnlich. Der Anteil kurzlebiger Arten ist auf allen drei Laubbaumarten relativ hoch und auch signifikant höher als auf Fichte (Die detaillierten Ergebnisse finden sich in Tabelle 2).
In Bezug auf die Geschwindigkeit der Wiederausbreitung ließen sich deutlich artspezifische Unterschiede feststellen. Ein Teil der Arten breitete sich unmittelbar nach dem Rückgang der S-Depositionen rasant aus, darunter zahlreiche Orthotrichum-Arten. Ander Arten, wie Porella platyphylla oder Frullania dilatata, die im Zeitraum 2005 bis 2008 nur selten gefunden wurden, sind nunmehr ebenfalls häufig. Schließlich besitzen aktuell auch weitere Arten punktuelle Vorkommen, welche 2005 bis 2008 keine epiphytischen Vorkommen aufwiesen; dazu zählen Ulota macrospora und Exsertotheca crispa.
Insgesamt wurden 119 Vorkommen von gefährdeten Arten nachgewiesen, darunter 30 Vorkommen von Orthotrichum pulchellum, 18 Vorkommen von O. rogeri, 12 Vorkommen von Cryphaea heteromalla oder 11 Vorkommen von O. scanicum. Mehr als die Hälfte der Arten besitzt keine historischen Fundorte in Sachsen, sondern wurde erst im 21. Jahrhundert erstmals dort nachgewiesen. Mit Ulota drummondii gelang sogar ein Wiederfund einer epiphytischen Moosart für Deutschland. Die Vorkommen gefährdeter Arten beschränken sich weitgehend auf die Laubbaumarten, wobei die höchsten mittleren Werte für Ahorn ermittelt werden konnten (s. Abb. 6).
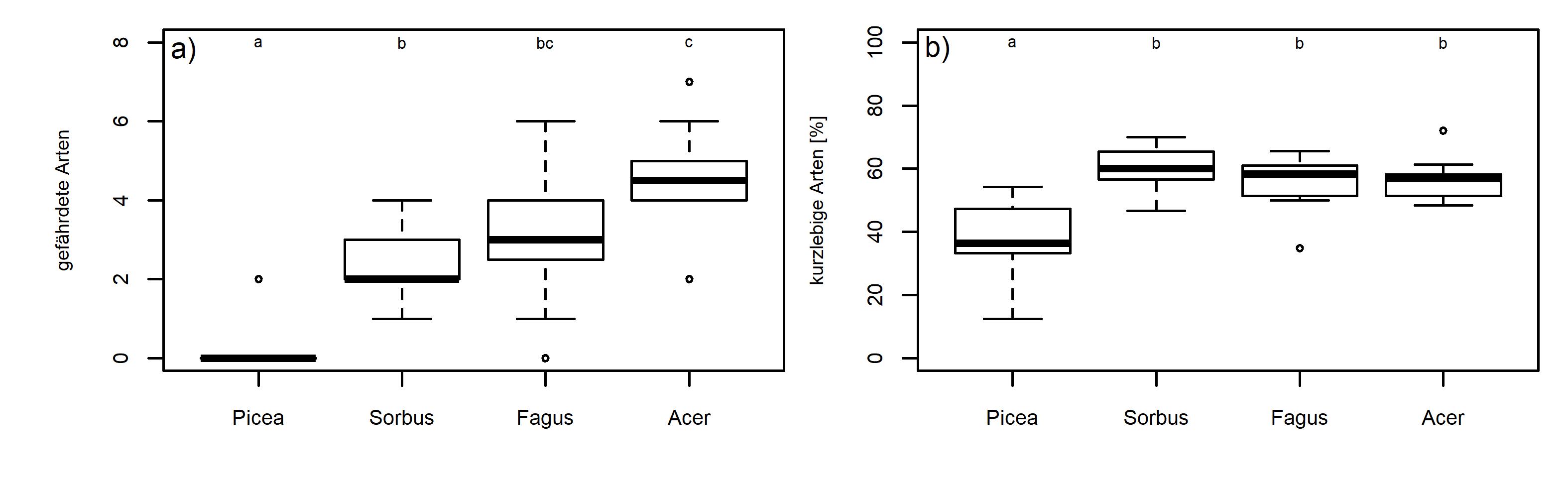
Abb. 6. Anzahl gefährdeter Arten (a) und Anteil kurzlebiger Arten (b) auf Probeflächen von Fichte, Eberesche, Buche und Ahorn, ungleiche Buchstaben indizieren signifikante Unterschiede (Dunn`s Post-hoc-Test nach positivem Ergebnis des Kruskal-Wallis Test, p ≤ 0.05). © Martin Baumann (SBS)
Was lässt sich daraus folgern?
Unmittelbar nach dem Rückgang der S-Deposition erfolgt auch in Gebieten mit vorher kritischer Belastungssituation eine rasante Wiederbesiedlung der Waldökosysteme durch epiphytische Moose, was die hohe Sensivität dieser Artengruppe in Bezug auf Stoffeinträge bestätigt. Da eine Regeneration versauerter Waldböden ohne Sanierungsmaßnahmen wie Kalkung nur über sehr lange Zeiträume stattfindet, zeigen andere systemare Indikatoren, wie die Krautschicht, keine dementsprechenden Reaktionen.
Nach einem Zeitraum der Wiederbesiedlung von ca. 15 Jahren repräsentiert die aktuelle Artenzusammensetzung epiphytischer Moose im Erzgebirge noch immer ein relativ frühes Sukzessionsstadium. Dies wird indiziert durch die Dominanz kurzlebiger Arten und eine erst allmählich einsetzende Wiederbesiedlung durch Arten mit limitierter Ausbreitungsfähigkeit wie beispielsweise Antitrichia curtipendula oder Neckera pennata. Insofern ist auch weiterhin mit einer hohen Dynamik hinsichtlich der Artenzusammensetzung zu rechnen. Dies wird besonders durch kontinuierliche Neu- und Wiederfunde ersichtlich. Seit dem Rückgang der S-Depositionen sind in Sachsen bislang Neufunde von 21 epiphytischen Moosen dokumentiert, was dem Trend in ganz Mitteleuropa entspricht.
Daraus lässt sich ableiten, dass sich die aktuellen Umweltbedingungen deutlich von denen zum Zeitpunkt vor Beginn der großflächig hohen S-Depositionen und dem dadurch verursachten Rückgang der Epiphyten unterscheiden. Diskutiert werden diesbezüglich insbesondere klimatische Veränderungen. Da viele der neu auftretenden epiphytischen Arten stickstoffreiche Standorte indizieren, lässt sich jedoch auch auf einen Einfluss hoher Stickstoffeinträge schließen.
Wie erwartet, ergaben sich auf den Laubbaumarten höhere Werte für den lokalen und den regionalen Artenpool, für die Anzahl obligater Epiphyten sowie für den Anteil gefährdeter Arten. Daraus lässt sich schlussfolgern, dass durch den Umbau von homogenen Fichtenbeständen in Bezug auf die Artengruppe der Epiphyten zum einen generell durch eine Etablierung von Mischbeständen eine höhere alpha-Diversität, zum anderen durch eine Anreicherung von Buche und Ahorn unter verschiedenen Standortbedingungen eine höhere gamma-Diversität resultiert. Die lange Zeitspanne der Sukzession und insbesondere die noch immer spärliche Verbreitung von Arten mit limitierter Ausbreitungsfähigkeit unterstreichen die Bedeutung einer Bewirtschaftungsform, die eine lange Kontinuität der Lebensbedingungen für die Epiphyten sichert. Eine Erhöhung der Diversität epiphytischer Moose in Fichtenbeständen lässt sich auch durch eine Beimischung der standorttypischen Eberesche (Pionierbaumart) erzielen. Im Kontext massiver Schäden in Fichtenbeständen und der Etablierung zahlreicher Sukzessionsflächen ist eine Beteiligung der Eberesche und weiterer Pionierbaumarten an der Baumartenzusammensetzung, abgesehen von der positiven Rückwirkung auf die Humusform, ein wesentlicher Baustein für die Entwicklung einer standorttypischen Biodiversität.
Weitere Informationen sowie Literaturhinweise finden sich im Originalartikel.



{kind=link}