
Over the past decade, there have been repeated extreme drought events in northern Bavarian forests, especially in Lower Franconia. These weather conditions have affected the water supply and thus the growth of trees. Second, they also reduced the forest’s resistance to abiotic damages and pests. Tree species that were once considered robust and vital, such as the European beech (Fagus sylvatica L.), showed signs of severe drought stress at many forest sites, especially in Lower Franconia. These ranged from premature leaf fall and crown damage to increased mortality (Figure 1).

Fig.1: Vitality loss of beech trees in Lower Franconia. Photo: Andreas Hahn (LWF)
This abiotic damage underscores the urgency of adapting to climate change in order to strengthen the resilience of forest stands. The search for resilient populations and their use in “assisted migration” (Fady et al., 2016) would be an important measure to increase the resilience of forests. However, moving a species within or even outside its range may disrupt ecological relationships and expose species to novel stresses. There is also the risk that introduced populations could become invasive, hybridize with local genotypes, alter community structure, or reduce local biodiversity (Aitken et al., 2013). We show, using the example of European beech, how forest damage survey data and precise observations on viability, vitality and water and nutrient supply can enhance the identification of resilient populations. This work was carried out as part of the sensFORnative project.
Field observations show that – contrary to most individuals of a population – some individual trees and specific groups of trees can maintain their vitality even under adverse environmental conditions. The influence of site-related factors on tree vitality is frequently observed and is considered ecophysiologically plausible.
The search for suitable seed stocks is based on visually and externally recognizable healthy tree populations, i.e. those that are apparently phenotypically resilient. It is obvious that observed resilience in populations could also be based on many factors other than genetic causes. Therefore, the search scheme focuses on genetically resilient populations by excluding other causes for the apparent resilience as far as possible, e.g. by taking site conditions into account (a high AWC is a key to bridge the water supply to trees during periods of drought; Marano et al., 2025). We demonstrate this here using a simple initial approach with a few parameters observed by us in this small preliminary study.
This simple search scheme is based on comparing the intensity of vitality loss and their vulnerability, which is determined by climate at the large scale and local site conditions at the small scale. The relationship between vitality loss and vulnerability can be represented graphically (Figure 2). Those forest stands or individuals that show only minor damage even during a drought period despite high site-related damage susceptibility can be expected to be particularly suitable under climate change conditions (Figure 2, green).
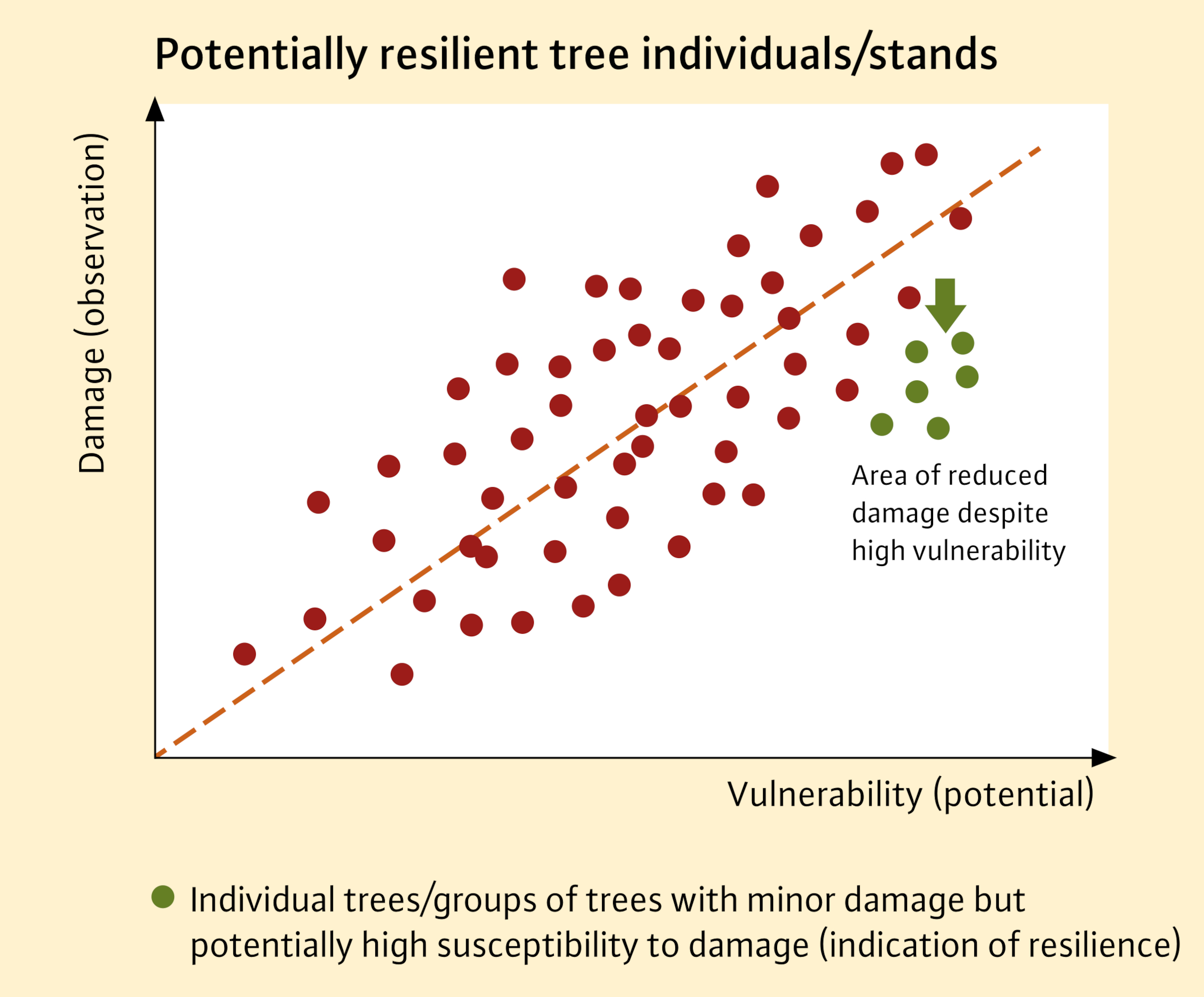
Fig. 2: Schematic representation of the relationship between vitality loss (observed damage) and potential site-related vulnerability/susceptibility to damage. The interaction of site characteristics such as water and nutrient supply, as well as the local climate, determines the potential susceptibility to damage of a site. Some forest stands may show below-average symptoms despite high susceptibility (green points below the red regression line).
Materials and Methods
Vitality loss and drought symptoms in beech are reported within the forest damage survey of the Bavarian Forest Administration (WSM reports). Data used for this study were recorded every six months between 2018 and 2021 for the total forest area (Schißlbauer et al., 2022). The forest damage survey includes all stands in a forest district; this means both damaged and non-damaged stands were considered. Beech stands had to show noticeable signs of dieback in the upper crown to be recorded, but the beech trees did not have to be completely dead.
The potential vulnerability of seed crop stands was determined in the sensFORclim project as climatic marginality (CMI) on the basis of niche models (Mellert et al., 2023). The tree species distribution data were taken from the Bayerische Landesanstalt für Wald und Forstwirtschaft (2020) dataset. In order to fully exploit the climatic potential of a tree species, beech was already considered present if it occurred within a 16 × 16 km grid. WorldClim (Fick & Hijmans, 2017) was used as climate data. The variables BIO6 (minimum temperature in the coldest month), BIO10 (average summer temperature), and BIO18 (summer precipitation) for the climate period from 1970 to 2000 were used as proxies for the limitations caused by chilliness and frost (BIO6), as well as for summer heat (BIO10) and drought (BIO18) (Mellert et al., 2015). For the large-scale representation of vulnerability, climate data for Bavaria were used, assuming a climate warming of +2.5 °C. This incremental scenario was used because such elevated Temperatures of this magnitude have already been observed in Bavaria in recent years. The resulting forecast of marginality (CMI) was mapped and classified using threshold values (CMI < 0.4 = marginal; CMI 0.4–0.7 = intermediate; CMI > 0.7 = optimal) to derive vulnerability (1 − CMI). On this basis, the CMI of European beech was mapped.
We then examined whether the classes of climatic vulnerability provide a meaningful classification. In addition, we tested whether there is a relationship between the CMI as a measure of potential vulnerability and the damage reports, following the concept described in Figure 2, using nonparametric correlation analysis with Kendall’s tau.
The data for the local site conditions include the defoliation rate of 332 trees located in twelve European beech stands in northern Bavaria (Schmied et al., 2024). The data, which were collected in the sensFORbeech project with a different objective (Schmied et al., 2024), are used here as an illustrative example of how the search process - from the stand level to the selection of plus trees - could be carried out (for further details, see Schmied et al., 2024).
Accordingly, the identification of “drought-resilient” populations relies solely on visual interpretation of deviations from the regression line; no statistical criteria were applied to identify significant outliers. The potential vulnerability of stands was described by the available water capacity (AWC) as a key driver of small-scale vulnerability (Marano et al., 2025; Mellert et al., 2018, 2023). This allows us to illustrate the relationship described in Figure 2 at the stand level, where AWC represents the potential vulnerability (x-axis) and the rate of defoliation represents vitality loss (y-axis).
Results and Discussion
The resulting area forecast of the CMI was divided into optimal, intermediate, and marginal ranges. A correlation between vulnerability and damage is plausible, but cannot be expected with certainty. This is because the niche models used are based on historical climate data (period 1970–2000) and a projected climate warming of +2.5 °C, while the WSM reports reflect current annual damage. This analysis can therefore be seen as a proof of concept to assess whether large-scale ecological parameters such as marginality, or derived variables such as vulnerability, are suitable measures of potential susceptibility to damage.
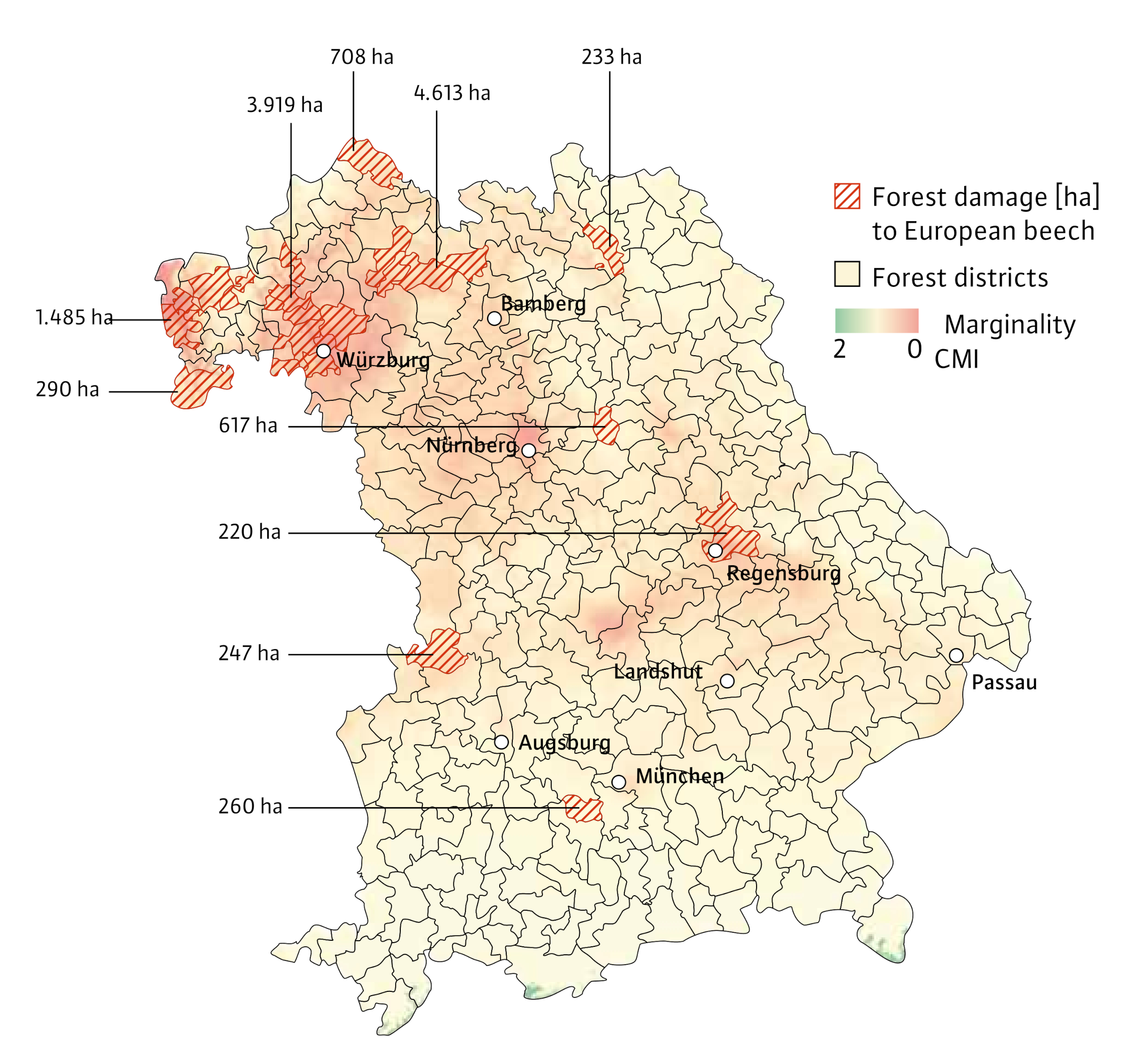
Fig. 3: Marginality of European beech in Bavaria (CMI) overlayed by the damaged areas reported for the forest districts of the Bavarian Forest Administration (regional offices for food, agriculture, and forestry) for the years 2018–2021 (hatched polygons).
As the map in Figure 3 shows, there is a correspondence between the CMI and the reported damage areas in the forest districts (Figure 4). Statistical analysis of the relationship between the two parameters provides more detailed information (Figure 4). The correlation between the two variables is significant (Kendall’s tau = –0.2; p < 0.001). The lower the CMI, i.e. the more marginal the stand, the greater the damage.
The scatter plot in Figure 5 shows that the greatest damage occurs in forest districts in Lower Franconia - a climatically highly vulnerable area (marginal area: CMI < 0.4). In the optimal range, on the other hand, less damage occurs (see Figure 3). These results suggest that the stratification method based on niche models is fundamentally suitable for assessing the climatic vulnerability of populations.
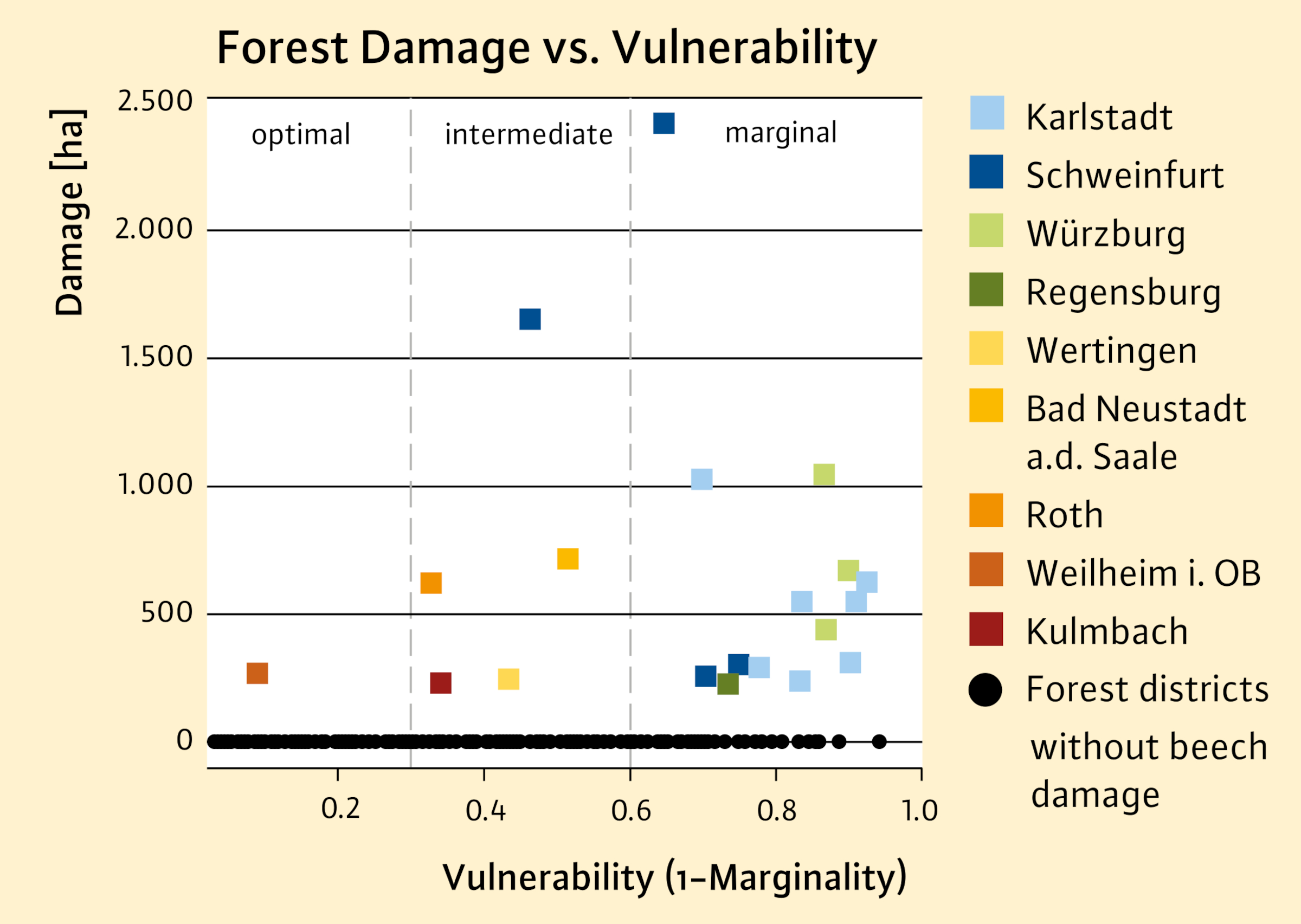
Fig. 4: Reported forest area with drought damage to European beech in Bavaria stratified according to vulnerability classes (V). Forest districts of the regional offices of the Bavarian Forest Administration with reported damages are represented by abbreviations. Particularly noteworthy are the forest districts of regional offices in Lower Franconia with high levels of damage (Karlstadt, Schweinfurt, Würzburg; see Figure 3). V < 0,3 = optimal; V 0,3 – 0,6 = intermediate; V > 0,6 = marginal.
The scatter plot in Figure 5 represents the effect of AWC as a driver of small-scale vulnerability on vitality loss (defoliation rate). Error bars in Figure 5 visualize the variation among individual trees within a forest stand. Increased climate plasticity is assumed in cases of below-average damage (stands below the dashed grey regression line). Promising trees or stands are those where high vulnerability is accompanied by only minor damage (highlighted error bars in shades of green).
According to our assessment scheme, three of the twelve tree populations stand out from the average trend, symbolized by the highlighted greenish error bars in Figure 5. These populations could therefore contain drought-resilient genotypes. We explicitly formulate this conclusion as a hypothesis for further testing. It is clear that an observed deviation from the expected damage does not in itself prove a genetic adaptation process. Therefore, the stands identified in this way should be examined in further studies and practical cultivation trials. Propagation material from stands tested in this way could make a valuable contribution to increasing the resilience and stability of forests.
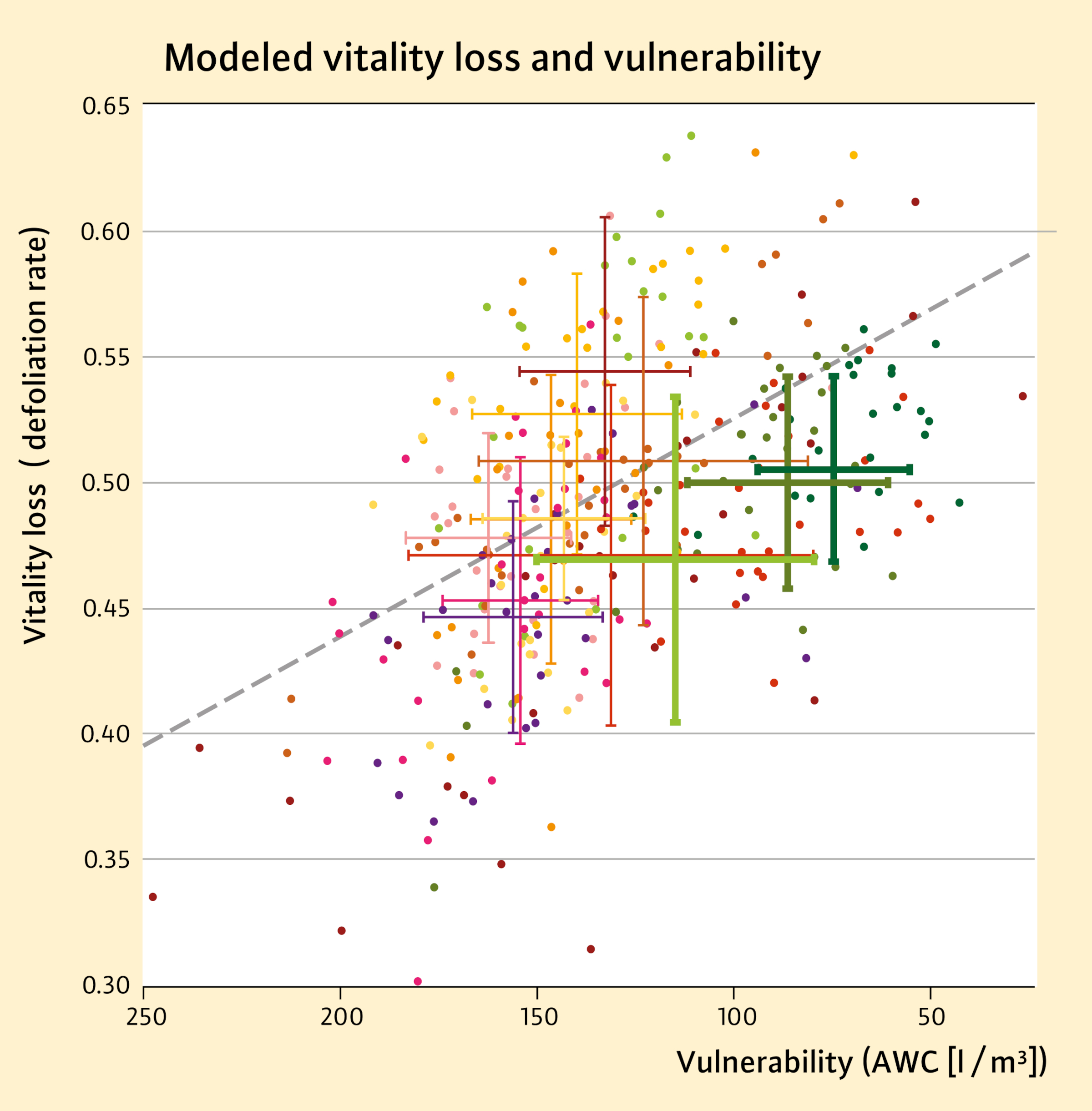
Fig. 5: Example for the identification of drought-resilient trees (N=332) within stands (N=12): Vitality loss (estimated degree of defoliation of individual European beeches in August 2022) vs. small-scale vulnerability represented by AWC in declining order versus increasing vulnerability.
Operational Procedure Despite Unresolved Issues

Fig. 6: Severe damage to beech trees due to drought in the main affected area in Lower Franconia. Photo: Yves-Daniel Hoffmann
After the severe drought events of recent years, the resistance and resilience of European beech have been questioned for large areas of forest sites (Klemmt et al., 2023). It remains unclear how these symptoms should be interpreted: Can the observed vitality loss and dieback be seen as an individual adaptation to new environmental conditions, or is a reassessment of suitable site requirements for European beech necessary?
Recent research in Bavaria, however, points to a complex response of trees to drought, depending on a variety of environmental conditions and spatial scales, including climate (Mellert et al., 2018), soil chemistry (Schmied et al., 2023), soil water storage (Schmied et al., 2024), and tree neighbourhood composition and structure (Rieder et al., 2026).
As shown here, relevant data on potential vulnerability (Mellert et al., 2023) and forest damage (Schißlbauer et al., 2022) are already available and can be used for a systematic search for climate-resilient provenances in Bavaria, even though the underlying processes of forest damage are not yet fully understood.
Nevertheless, the search procedure described here needs to be further developed using more comprehensive datasets. Future application-oriented research should include forest protection data (covering all types of forest ownership and longer time series), data from forest condition surveys, updated site information (Mette et al., 2025), and high-resolution monitoring of damage symptoms using remote sensing. Initial studies already provide reason for cautious optimism (Pfenninger et al., 2025).
Conclusion
Damage surveys at the forest district level provide a good overview of the distribution of biotic and abiotic damage in forest stands; hotspot regions can be identified. The correlation between forest damage and climatic marginality, which serves as a criterion for the regional susceptibility of forests to damage, highlights the important role of macroclimatic drought risk.
Climatic marginality can therefore serve as a starting point for a systematic search for drought-resilient populations. Stands with little or no vitality loss in such problem areas may indicate higher resilience. Such phenotypically promising candidates, identified through observational classification, can be included in a systematic testing procedure.
In particular, site-specific causes for (apparently) increased resistance should be excluded as far as possible. A tree population identified as more resilient in this way could - after thorough testing in accordance with the German Forest Reproductive Material Act (BLE, 2003), for example with regard to quality and genetic adaptability - be designated as a seed stand for drought-resilient propagation material.