
The grey larch budmoth (Zeiraphera griseana) occurs all over Central and Northern Europe and its range extends as far east as Siberia. It is a small, speckled grey-brown moth with a 2 cm wingspan (Fig. 1).

Fig. 1. The grey larch budmoth. Photo: Beat Wermelinger (WSL)
Its larvae reach a length of 1.5 cm and occur in a range of colours. The so-called larch form (for more information see below) has a black head capsule and is initially light in colour but later turns grey-black, whereas larvae of the yellowish-grey cembran pine form have an orange-yellow head capsule (Fig. 2).

Fig. 2. The larch form’s black-grey caterpillars (left) feed almost exclusively on larch needles. The yellowish-grey caterpillars of the cembran pine form (right) mainly feed on the new shoots of cembran pine, but they also occur on larch, Scots pine and spruce. Photos: Beat Wermelinger and Beat Forster (WSL)
The larch moth has a one-year life cycle. The young larvae hatch in mid-May and pass through several larval stages. For its development a single larva needs between 10 and 20 clusters of needles, but it eats just half their needle mass (so-called wasteful feeding). At the end of their roughly four-week development, the larvae lower themselves to the ground and pupate in the litter. The moths emerge from the end of July to September, swarming between dusk and midnight. Mating takes place during these flights, and the females then lay up to 300 eggs.
The population cycles
In lower elevations of the Alps and on the Swiss Plateau, larch buthmoth populations always remain small and thus inconspicuous. Only in higher valleys with a more continentally influenced climate do mass outbreaks occur at more or less regular intervals. An intensive study of the population dynamics of the larch budmoth, focusing on the Upper Engadine, was started in 1949 (Fig. 3).
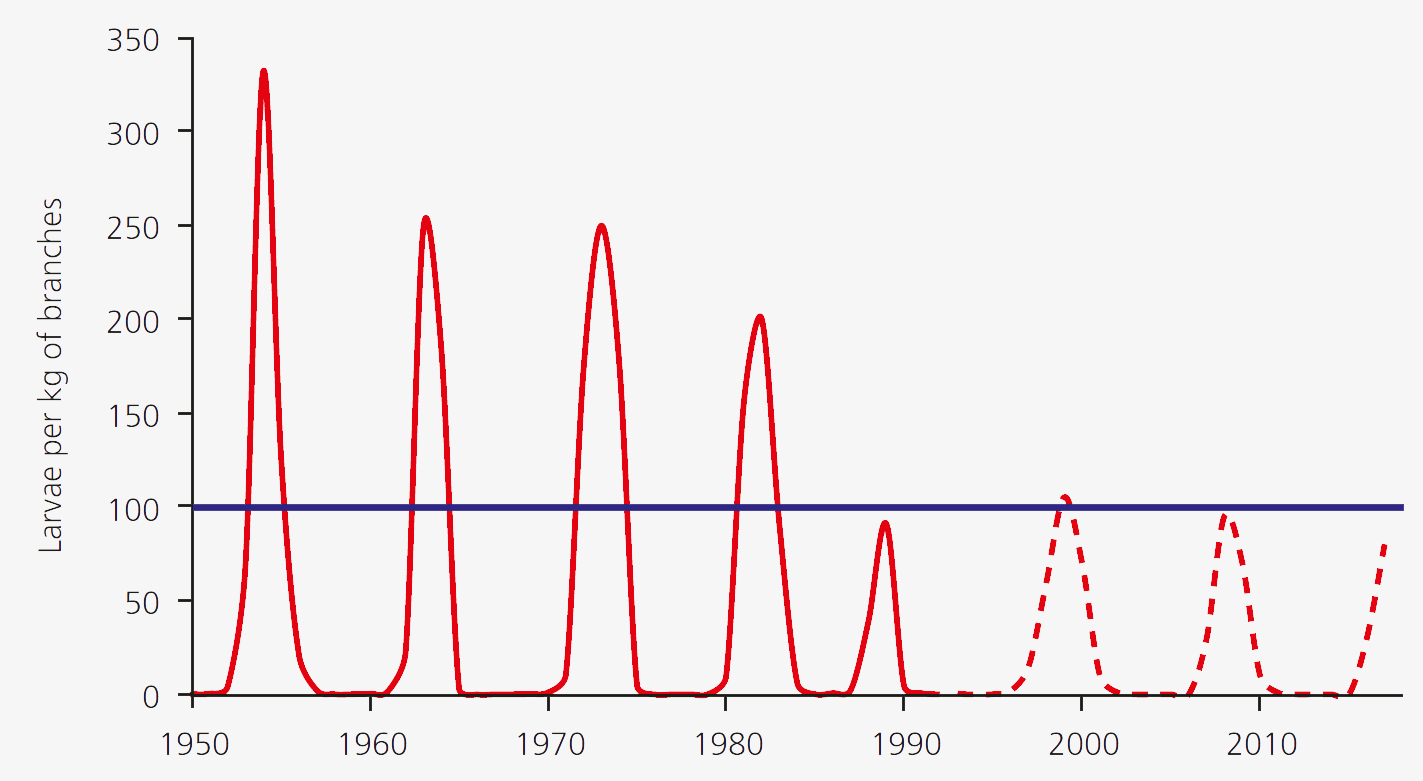
Fig. 3. Cyclical population fluctuations of the larch moth in the Upper Engadine. The mean periodicity is 8.5 years and the threshold for visible discoloration of larch stands is 100 larvae per kilogram of branches (blue line). The solid red curve is based on data from Baltensweiler (1993a) and Baltensweiler and Rubli (1999). The dashed line is based on visual estimates made in the field.
It emerged that outbreaks tend to occur, on average, every 8.5 years. Within four to five generations, population density fluctuates by a factor of up to 30,000! At the peak of an outbreak (Fig. 4), there are more than 20,000 larvae on a single larch tree, and the sound of their faeces falling to the ground is audible.

Fig. 4. At the culmination of the roughly nine-year cycle, larch trees infested with larch budmoth larvae turn reddish brown. The cembran pines in the foreground are unaffected. Photo: Beat Wermelinger (WSL)
The discoloration of larch stands only becomes visible at a threshold of about 100 larvae per kilogram of branches and with 10% of the needles being damaged. By the mid-1980s this threshold was regularly being vastly exceeded. The regular cycles in the Engadine are limited to the optimum larch budmoth habitats, situated between 1,700 and 2,000 m a.s.l., and to forests with a sufficiently large proportion of larch trees. Similar cycles occur across the entire Alpine arc, from the French Maritime Alps to Carinthia in Austria.
The larch growth in years when infestations occur is severely stunted, and this is reflected in narrower tree rings in their stems (Fig. 5). Dendroecological studies have shown that larch budmoth cycles have occured for a very long time. Wood samples taken from living larch trees and from beams in historical buildings were used to reconstruct larch budmoth cycles spanning 1,200 years in Valais.
A budmoth population builds up and collapses again in just a few generations. During a collapse, the larval mortality rate owing to antagonists, competition and starvation peaks at 99.98%.

Fig. 5. Outbreaks of the larch budmoth are often visible in the regularly recurring, narrow annual tree rings in the stems of older larch trunks. Photo: Beat Wermelinger (WSL)
What governs larch budmoth cycles?
The astonishing regularity of larch budmoth cycles begs the question of the mechanisms that cause them. There are various processes that take place during mass outbreak and shape the cycles:
The negative feedback of needle quality
Larval development is closely related to larch needle quality. The loss of green needle mass means larch trees produce a smaller amount of photosynthates for their growth and reserves. If they lose more than half their needle mass, they bud again at the end of July, producing a second set of needles. However, they do so at the expense of their reserves. Their needles grow more slowly the following spring and remain between 30 and 70% shorter. The needles additionally have a higher raw fibre content and tend to contain less protein and nitrogen than in years without mass outbreak.
As with all leaf-feeding caterpillars that overwinter at the egg stage, the timing or degree of synchrony between needle budding and the emergence of the young larch budmoth larvae is crucial. In the years after a mass outbreak, the synchrony of these events worsens and larvae emerging at the usual temperature-triggered time miss fresh, tender, readily digestible larch needles. As a result, at least the earliest larch budmoth larvae of the new population end up starving. Those larvae that do survive find needles of inferior quality that are difficult to digest and have a lower nutritional value. As a result, they grow more slowly, gain less weight and have a higher mortality rate. Moreover, the moths that develop from these larvae produce up to 90% fewer eggs. This delayed negative feedback triggers a drastic decline in the moth population density.
Regulation by natural enemies
The most important driving force are wasps and flies parasitizing the larch budmoth larvae. The cycles of these natural enemies lag about two years behind those of the larch budmoth itself. At the start of an outbreak, the mortality rate among larch budmoth larvae caused by parasitoids is mostly less than 10%, rises to 20% at the culmination of the outbreak, and is as high as 80% when the population collapses.
Fitness of different ecotypes
The larch budmoth has two genetically different ecotypes (varieties): the larch form and the cembran pine form (Fig. 2). The larch form’s grey-black caterpillars feed almost exclusively on larch needles and exhibit a very high mortality rate on cembran pine. Its eggs develop significantly faster and larval mortality on high-quality larch needles is lower than for the cembran pine form. The yellowish-grey caterpillars of the cembran pine form mainly feed on new shoots of cembran pine, but they also occur on larch, Scots pine and spruce. The cembran pine form develops much more slowly and produces fewer eggs than its larch counterpart, but survives better than the larch form on poor-quality larch needles.
At the onset of a mass outbreak, the darker larch form predominates at 80%. After complete defoliation at the culmination of an outbreak, budburst of the larch trees is delayed and they produce needles of inferior quality. At this point, the later-emerging cembran pine and intermediate forms, which survive better than the larch form on such needles, have an advantage. Only when needle quality has improved does the fitter larch form prevail again and the reproduction rate of the population increases once more.
Moth migration and dispersal
If complete defoliation occurs in ideal habitats, subsequent generations of moths move on to lower-lying areas, where the larch trees were spared excessive larval feeding, to lay their eggs. Once the trees have recovered, moths fly to higher elevations over the next few years in the evenings against prevailing valley winds, thus returning to the optimum habitats characterised by better synchrony.
At a larger spatial scale, the windborne dispersal of larch budmoths between inner Alpine valleys is important. In areas where larch trees have been largely defoliated by an outbreak, moths leave in large numbers and are blown into neighbouring valleys by the prevailing westerly wind.
Climate change
The maximum population densities during the last three cycles (1989, 1999, 2009) did not reach even half the previous values in the Engadine (Fig. 3), which is why in most places the larch forests were spared widespread discolouration. In France as well, far lower maxima were reached during this period. The reasons for lower maxima in recent cycles are unclear but are probably related to rising temperatures over the last few decades.
Warmer autumns and milder winters increase the respiration of overwintering eggs which could cause their energy reserves to become exhausted prematurely. More importantly, the crucial synchrony between larval hatching and the emergence of needles may have worsened in the spring. If larvae starve because they hatch before the needles emerge, this has a major impact on population density.
The importance of the larch budmoth
Whereas older literature describes the larch budmoth as a real pest, intensive research has significantly relativized such a negative categorisation. Studies in the Upper Engadine have shown that fewer than 1% of larch individuals usually die because of an outbreak and that their loss of growth is negligible.
Lengthy tree-ring series show that larch trees and the larch budmoth have co-existed for millennia. Larch trees are highly dominant in many places today because, as a pioneer species, they have been able to benefit from previous disturbances. For instance, in the Middle Ages many spruce/cembran pine forests were cut down, the war years were characterised by conflagrations, and cembran pine has been deliberately targeted for use as wood panelling since time immemorial. In addition, up until the end of the 19th century the regeneration of these two tree species suffered as a result of wood pasture and mulching. These human activities enabled larch, a fastgrowing, light-demanding, pioneer tree species, to establish a firm foothold. However, a dense, old larch forest barely regenerates, and shade-tolerant species such as cembran pine and spruce establish and ultimately form a climax forest in the absence of any new disturbances.

Fig. 6. Val Guisane in the French Alps. Although the infested forests look "sick" to the layman, this millennia-old phenomenon is a natural process in the forest dynamics of larch/cembran forests. In most cases, only a few trees die as a result. Photo: Forest Entomology WSL
While larch trees can compensate for being stripped of their needles by rebudding, cembran pine can only do so to a very limited extent, so any affected parts of their crowns die back. Larch trees can then capitalise on the death of cembran pine individuals, making use of newly available space and a greater incidence of light. These processes limit the development of cembran pine trees in the optimum larch budmoth habitats and thus delay the transition of larch forests to mixed larch and pine climax forests. Consequently, the larch budmoth is of great ecological importance to inner-Alpine larch forests.
Control measures
When larch budmoth larvae feed on larch trees, the negative effects remain minimal and hence do not justify the implementation of any control measures. During heavy infestations, it is important to promptly explain to the local population and holiday guests why the forests are turning brown and to promote an understanding of this spectacular phenomenon.
Literature
Literature references used in the text can be found in the WSL fact sheet Cycles and importance of the larch budmoth. You can order the printed publication free of charge from WSL:
WSL e-shop
Zürcherstrasse 111
CH-8903 Birmensdorf
e-shop@wsl.ch